|
1. Muscles →> cartilage
→> bone, tissue types:
Muscles get encapsulated in connective tissue and tendons of connective
tissue attach to the skeleton of cartilage or bone.
Cartilage and bone originates like muscles from
mesoderm, the 3rd layer in the gastrulation of the embryo.
It's characterized by scattered cells in extracellular fiber substance,
corresponding to step 2 - 1 in the suggested chains of tissue types
(file Levels, No.
5).
There are several features besides the tissue
type that support the view on skeleton as characterized by dimension
degree (d-degree) step 2 - 1 in a dimension
chain of organs - to the extent that the organs can be interpreted
as such a chain. It's also characterized by anticenter in several
ways:
- The mentioned origin from mesenchyme cells, i.e. free cells at
the end of the level chain.
(Bone can also derive from cartilage.)
- Osteoblasts and osteocytes are formed from the neural wall (Kz
p. 16) around the neural plate, original animal pole 00
of the embryo.
- Bone cells become oriented in concentric circles in the tubular
bones as anticenter around blood vessels.
- Inorganic material is built-in into the skeleton from the surroundings,
metal ions as Ca2+ for instance (metals
representing the 00-pole in relation to non metals among elements
in the periodic
system). Notice that the same concerns blood, the last,
5th kind of tissue. Metal ions from surrounding anticenter become
part of the structure, not only free ions in liquids.
- Elementary forms of skeleton on a macro-level is "1-dimensional"
needles or tubes, straight or bowed, or 2-dimensional "surfaces",
bone-plates or shells.
An exoskeleton is clearly a surface, d-degree
2. In an endoskeleton as supportive structure the spinal column
and skeleton of limbs give the basic form of the organism as lines
(d-degree 1) like elements in an architectural drawing.
2. Evolution:
Already unicellular organisms show elementary forms of skeletons
as spicules or shells. Skeleton substance get stored as layer outside
the epithelium of the skin, for instance by corals.
One-layer organisms as sponges produce needles,
spicules in the substance inside the outer epithelium, between outside
and inside - as between d-degree poles 2a and 2b (!):

Fig Sk-1-57
Spicules of the sponges have a lot of varying shapes,
"drawing elements" that differentiate species: with /
without endplates, S-formed, bowed with opposite directed, concave
and lobated endplates, radial three-numbered or 6-numbered with
secondary ramifications etc. They seem to express geometries of
d-degrees 1- 2 - 3. (Radial /circular
shapes for instance, poles 3b - 3a in our model, are outer poles
of d-degree 2.)
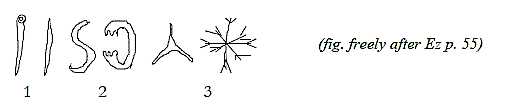
Fig Sk-2-58
It would be very interesting to know how these different shapes
are expressed in genetic differences!
A main division in evolution of species among
3-layer animals is the one between Protostomia as arthropods
with a skeleton as an outer shell, an exoskeleton, and Deuterostomia
with endoskeleton: a spinal column in vertebrates and many staff-
and tube-shaped elements.
This opposition concerns origin of material from
ecto- versus endoderm, thus directions from outside or inside respectively,
poles 4a and 4 b in the model, which also are assumed as "outer
poles"of d-degree 3. See further Evolution.
It shows also features of the polarity circular - radial structures
in next step 3 - 2.
A secondary polarity of tube-shaped bones is the
one of convex versus concave ends, geometries of poles 2a and 2b
of d-degree 1 (the tubes on a macros-scale):
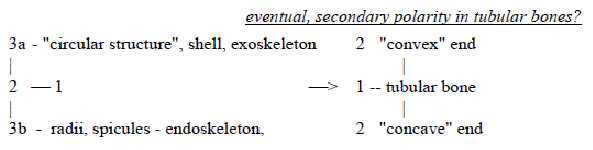
Fig Sk-3-58-2
However, also the vertebrates develop shell-like plates as
around the brain and in the pelvis. There are no absolute borders.
The primarily most massive, "3-dimensional"
skeleton element in vertebrates seems to be the vertebra. It could
be connected with the fact that they in evolution develop as bony
replacement of the notochord, which originate from archenteron and
the vegetative pole, the endoderm.
3. Skeleton of the trunk in vertebrates:
In a lancelet, regarded as a basal subphylum of chordates, the
skeleton of the trunk as a whole gets roughly the same structure
as the blood system (lacking blood cells). (Note that both
make up the last tissue types where also metal ions are built in
as elements of structure.)
The notochord stretches along dorsal aorta, ribs
along arteries of gills, breastbone along ventral aorta.
 Fig
Sk-4-58-3 Fig
Sk-4-58-3
Simultaneously, the relation blood system - skeleton shows opposite
directions: ribs depart from the dorsal side (~ 00-pole), from the
spine, and only some of them reach the breast bone. While the blood
stream through gills has opposite direction, goes from ventral aorta
(~ 0-pole) upwards through gills towards the dorsal one. It's a
polarity as from V- and A-poles respectively, ventral - dorsal sides,
originally from inside - outside.
(It may also be noted that the ribs on the ventral
side bow upwards towards the front part: ventral side as from 0-pole
with direction forwards seems to have an impact on this secondary
feature.)
The ribs - in the plane of the 3rd coordinate axis right-left (from
step 3 - 2 in the embryo development) get roughly a circular structure.
The segmentation of the ribs becomes analogous to the segmentation
of somites of the embryo, which gives striated muscles, i.e. characterizes
front and dorsal parts as polarizations from the 00-pole.
 Fig
Sk-5-59-1 Fig
Sk-5-59-1
The complementary features in dorsal - ventral design appear
in later chordates in the surface character of the shoulder blades
as rest of a dorsal ("circular" ) shield
versus the radial character of collarbones (clavicles) on
the ventral side; the opposition 3a - 3b in he dimension model.
It could be added that the ribs are 2 x 12, at least in humans,
as sum of 5 + 4 + 3: cf. the genetic
code and 14 and 10 in the 2x2-series behind
the periodic
system. The 12 pairs divided: 7 (attached to breastbone),
5 divided into 3 that are fused and join the 7, + 2 "floating"
without any ventral contact.
Shoulder blades and bones of the pelvis
with ridge and "knobs" could in themselves illustrate
forms of d-degrees 2-1-0. Pelvis bones of back part (from vegetative
pole) form more of a circular form from d-degree 3, and are bigger
in size.
The bones that cover parts of the dorsal side
underlines skeleton as an organ characterized by anticenter: distal
side representing original 00-pole. They serve as shelters but also
as attachments for muscle tendons of the limbs, the "extremities".
About the production of blood (from file Blood):
Blood is at first in the embryo and among early chordates produced
by the spleen, a gland developed from the vegetative and nutrition
system. Then it becomes produced in the skeleton.
This "changeover" could be interpreted
in accordance with the loop version of the dimension model, where
the d-degree step 4 →>3
correlates with step 2←1
inwards the nutrition system. a turn in direction towards the nutrition
system to which blood system belongs.

Fig Sk-6-59-2
4. Why extremities and why from surface of the body?
It feels natural (at least for a closer relative to a starfish)
to think of extremities as radiating from a center, bones in extremities
departing from spinal column or the like. This is however not the
case. It's truer about blood and muscles but skeleton for arms and
legs are derives from lateral tissue at the surface of the body
like the spicules of sponges mentioned above, and they are only
loosely connected with skeleton of the trunk. A rather curious fact.
(Cf. on the unicellular level where basal bodies of cilia
are positioned right below the surface of the cell membrane.)
One simple aspect on this fact is the simple geometry on a macro-scale:
Trunk →>
Skin →> limbs as d-degrees
3 → 2 →
1.
The same simple view could be applied on ciliated unicellular organisms.
(However, we should eventually see d-degree 1
in this case developed "between" trunk and skin in step
3 - 2; cf. that 3 of the 5 origins of arm muscles in humans, serving
the elbow joint, attach to the trunk, 2 to the forearm.)
Another aspect is function: the limbs become vectors, developed
as the fundamental, dimensional polarity 0 == 00 of d-degree 4 between
whole organism as 0-pole, defined by its demarcating
skin, and its surrounding as 00-pole (water or later solid
ground).
The extremities serve locomotion, d-degree 0/00
of the dimension model, locomotion in the external world.
Tubular bones of limbs develop and get strengthened through pressure.
Pressure (F/m2) becomes
a 2-dimensional force when the primary force as in the model here
is defined as a 4-dimensional vector (e.g. FG,
gravitation). Hence, the limbs could be suspected to be primarily
straight (linear) vectors as counterforces to this pressure.
(In this sense legs could be seen as created or
induced from outside, from the environment, with a divergence force
from the pressure pointing inwards the body and radiating to several
(5) segments of spinal cord for innervation.)
However, the development of legs in 3-layer animals must have a
deeper cause. Internally given. Segmented animals as e.g. millipedes
have 1 pair of legs on each segment, segments that in human beings
are merged and united to 2, head uncounted. (Already the division
of mesoderm
shows this division into 2 differing "segments".)
Segments get obviously uniting during evolution.
(Cf. that 5 foremost segments of neural tube in crabs become its
brain.)
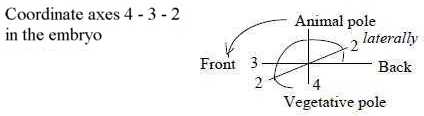
Legs develop along the lateral axis left-right, L-R (as in crocodiles),
and become later among land living chordates mostly or partly parallel
to the dorsal - ventral axis, which corresponds to original animal
- vegetative axis (A-V). With first 3 axes, A-V, F-B and L-R representing
d-degree 4 - 3 - 2 (see file Embryology),
legs appear given as the 4th axis, corresponding to d-degree 1.
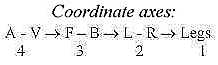 Fig
Sk-7 Fig
Sk-7
Simultaneously, while legs of chordates develop from the paired
(hence "lateral") fins of fishes, many fishes have also
dorsal and ventral fins. Cilia →>
bristles →> fin rays are
to regard as the general principle of something sticking out as
vectors or "field lines" into the surrounding, getting
many functions for both sensations and locomotion.
5. Tetrapods and their extremities:
On a macro-scale a tetrapod gives a picture of a pentagon or hexagon
(with or without tail!), as the rings of the molecules in RNA -
DNA or in carbohydrates as riboses - hexoses. (Leonardo da Vinci,
who drew the man in a circle, didn't know about molecules and their
forms!)
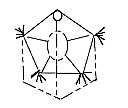 Fig
Sk-8-60-1 Fig
Sk-8-60-1  Fig Sk-9-60-2
Fig Sk-9-60-2
Number 4 in extremities:
Number 3 in division of the body: head, front part and back part
of the trunk.
Number 5:
Why is exactly this number 5 developed in limbs of chordates? In
human hands and feet? Hardly accidental. There is the same 5 in
"5-merous"
plants. Cf. also development of the 5 brain bladders. Number
of gill arches is also usually 5 in cartilaginous fishes (Kz
p. 29). And as said above there are the muscles with 5 origins
to the arm, nerves from 5 segments of spinal cord (vertebra) to
arm etc.
(It is said that the original segmentation of
somites, of muscles etc. among fishes has been indistinct in tetrapods
but can be seen in the embryos (Kz p. 179).)
The 4-legged fish Ichtyostega, found on Greenland, had already
the 5-rayed design and all the 27 bones of a limb including hands,
forearm and upper arm bones of a developed land living chordate.
The number 5 appears already among the invertebrates,
early precursors to the vertebrates, as for instance in the number
of arms of starfishes, although secondarily developed. Among invertebrates
along the other line of evolution there is the big group of decapods
among crustaceans with 5 pairs of extremities, and further for instance
the ten-armed cuttlefish (cephalopod) with 10 tentacles.
Pairs of segments that get used as legs: either
a multitude or → 4 →
3 → 2→
1: crustaceans 4 where the 5th pair becomes gripping (prehensile)
claws; spiders 4 pairs; insects 3 pairs, mammals 2 pairs, birds
and human beings 1 pair.
Here is this number 5 naturally assumed as rooted in the 5-dimensional
background model, a development of number of steps in a dimension
chain:
Transformation of a dimension chain as structures to number of
something:
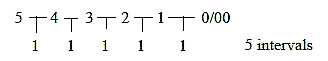 Fig
10 Fig
10
6. Geometrical forms of arms and legs as a dimension chain:
D-degrees of geometry: Finger tips 0/00: points;
↑ hard nails (dorsal side ~ 00), soft tips (ventral
side ~ 0)
↑ Fingers: 1, lines
↑ Palms: 2, surfaces
↑ Wrist bones: 3, "nodules",
~ mass
↑ Arm bones: 4, vectors: outwards
1, "inwards" ← 2.
Joint types, 3 →>
2 →> 1 bending axes:
Types of synovial joints in the body are about 5 or 6 if we include
the gliding ones between metacarpal bones: Ball and socket j., Pivot
j., Ellipsoid (condyloid) j., Saddle j., Hinge j., Gliding j. (http://www.teachpe.com/anatomy/joints.php).
There is mainly decreasing degree of flexibility from shoulders
and hips towards fingers: To simplify:
- Shoulders and hips: Ball- and socket joints, a 3-dimensional motion.
- Elbows and knees: Hinge joints + the pivot, rotation type between
radius and ulna in forearm and corresponding bones in calves. Motions
in 2 planes.
- Wrist: Condyloid (ellipsoid) joints between forearm and carpal
bones: More limited motions in 1 of 2 planes.
- Saddle joint of the thumb, allowing rotation in 1 plane.
- Finger joints between phalanges: hinge joints, mobility in only
1 plane, extension / flexion.
The directions of mobility get more crystallized towards one-way
direction outwards the fingers in accordance with a main view in
the dimension model. Simultaneously, the "freedom of degree"
increases naturally with every joint along the limbs - as d-degree
of motions increases towards lower d-degree of structure in the
model. (However in about 7 steps in tetrapods).
An outline of the directions of movements as angle steps:

Fig Sk-11-62-1
7. Hands and feet:
- From wrist to fingertips (radially) there are 6 bones, proximal
and distal wrist bones (carpal bones in approximately two rows),
palm bones (metacarpal bones) + 3 finger bones (phalanges),
2 in thumbs.
- Left-right (circular) there is principally 5 branches.
Cf. 6 "borders", 5 intervals in the dimension chain:
 Fig Sk-12-62-2
Fig Sk-12-62-2
A certain differentiation appears in human hands also among the
"branches", here illustrated as angle steps of a dimension
chain:
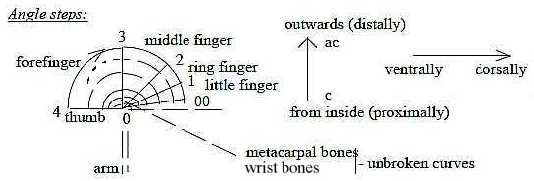
Fig Sk-13-63-1
(Cf. Some of the names: "forefinger", in Swedish "pekfinger"
= the pointing finger, is associated with direction, d-degree 4.)
Tendons:
Details of tendons in the hand and its innervation can be interpreted
with aspects from the dimension chain.
Relation between ventral palm and dorsal back
of the hand originates from the 0- and 00 polarity which also is
a relation between higher and next lower d-degree. The loop version
of the model implies 3 polarizations of 5: 5 →>
0 - 00, 5' →> 4 - 1, 5"
→> 3 - 2.
Dorsal side (~ 00-pole) of fingers has no muscles,
only tendons, connective tissue of lower d-degree than muscles according
to the identifications here.
While ventral side of fingers and the palm (~ 0-pole) includes muscles.
Tendons to fingers, number division:
- Back of hand, dorsal side: division to fingers: 1 - 3 - 1
- Palm, ventral side: division to fingers 1 - 4.
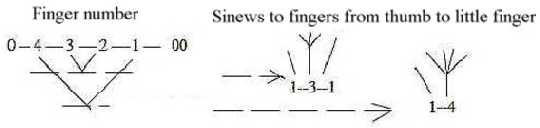
Fig Sk-14-63-3
Innervation:
The innervation - if rightly understood here - follows a similar
scheme of divisions 4-1 to 3 - 2 when it concerns the polarity motor
- sensory nerves, outward/inward direction,
also a polarity corresponding to higher versus lower d-degrees:
The motor nerves:
- N Medianus through the center of forearm and wrist innervates
the bigger muscles of the thumb and its opposition to the other
fingers.
- N Ulnaris passes through forearm more distally and innervates
the smaller muscles of the other 4 fingers plus their lateral movements.
Hence, the motor innervation divides the fingers
1 - 4: thumb - the other ones.
The tactile, sensory innervation divides them approximately
3 - 2 (or 3.5 - 1.5):
- N Medianus innervates the 3 (3,5) inner fingers,
- N Ulnaris the 2 (1,5) outer ones.
The arrangements illustrate how the opposite directions outwards
→ ← inwards (~ poles
4b - 4a) between motor and sensory nerves also simultaneously appear
as a d-degree step in number division 4 - 1 to 3 - 2, in accordance
with the loop version of the dimension model.
It's notable that the sensory innervation as inward
direction from anticenter also is displaced a bit, half a step towards
the 00-pole ~ outer side.

Fig Sk-15-64-1
Carpal bones 8 (+/-1):
There are 8 bones lengthwise from shoulder joint to finger tips
and 8 (+/-1) carpal bones arranged
as in two rows along a transversal axis, a turn 90° The number
could eventually be interpreted as the sum of vectors poles 4b and
4a, turned to a 90° of polarity in d-degree 3, see figure below?
Another aspect concerns the transformation between
numbers 5 and 8: how 5 become 8 or 8 becomes 5 - a principal sketch:
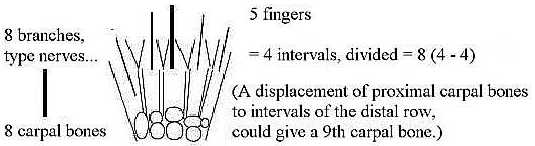
Fig Sk-16-64-2
The arm lengthwise = 8 steps, interpreted as 5 in
accordance with the same principle:
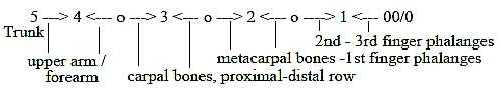
Fig Sk-17-64-3
Compare the interpretation of geometrical forms
above:
- upper arm-forearm as vectors (4) outwards /
"inwards"*,
- wrist bones (carpal bones) as nodules (3),
- palm bones (metacarpal bones) as surface (2), cf.
web between fingers / toes !
- finger phalanges (1).
How to motivate implications of such a figure?
- That bones principally are formed through the opposite forces
outwards / inward pressure may not be
too difficult to imagine.
- Tube-formed bones have more or less marked one convex and one
concave end - a polarity of d-degree 2 in the model and in agreement
with our interpretation above of skeleton as organ of d-degree step
2-1 in a dimension chain of organs.
In addition there is the polarity in thickness
between the 2 bones in forearms and calves. (The number relation
2 - 1 between bones of forearm and upper arm could also be seen
as expression for the inward relative outward direction.)
The change at the wrist, in the figure in step 4→>3,
is principally an angle step from 180° to 90° as assumed
in the dimension chain: the limb transforms as induced from outside
to a surface (d-degree 2) characterized by circular and radial structure
in accordance with outer poles of d-degree 2 from d-degree 3 in
the dimension chain ("circular" in arrangement of carpal
bones in the wrist, in demarcation of the palm outwards - and perhaps
also later in the opposition of thumb to other fingers).
The impact of meeting the environment as anticenter
and 00-pole can surely be seen in the transition:←2 ← 1←
00.
In other respects, as divergence from each other joint and convergence
to the other joints, the figure seems difficult to motivate.
[Arm versus hand reminds a bit of the double nature of light
as particles and waves: the wave patterns that appear when photons
pass through holes of a certain small size in a screen. What
could have served as the "hole" for passage when "straight"
forearm transforms to 8 (-/+1) carpal
bones and 5 "waves"? The contact point with ground
a reasonable answer?

Fig Sk-18-65-1 ]
8. Reduction of toes:
The big toe, the most proximal, disappears in tetrapods of the
kinds below - as a displacement outwards in distal direction. In
the history of evolution the even-toed mammals are later than the
odd-toed ones (Fz p. 147), also a displacement half a step
outwards. As illustrated here, a displacement of the middle:
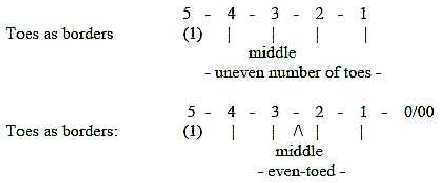 Fig
Sk-19-65-2 Fig
Sk-19-65-2
Cf. Evolution:
the direction of evolution towards Deuterostomia and chordates as
inwards higher d-degrees. The 5th toe, the big one, and thumb could
be regarded in the same inward direction as first fully developed
in use with the primates.
9. Number of bones in human extremities:
Bones in hands are 27, divided: 8 wrist bones, 19 finger bones.
We can observe that it is numbers in the x3-series (x
= 5-0) in step 3 - 2:
x3: 125 —
64 — 27 — (19) — 8
— 2 — 0
Adding 3 bones in the arm plus scapula and collar bone it makes
32, 1/2 x 43.
32 is approximately the same number as vertebra in the human spinal
column.
Legs: 3 pelvic bones, 3 bones in the leg, 7 ankle bones and 19 toe
bones gives also 32.
* 7 bones in human ankles make 26 in feet. However, wrist and
ankle bones in original type of tetrapods are said to be 9 according
to one source (Kz p. 84).
Number of bones in a human being approximately around 216 = 63.
The information can vary, probably depending on if some bones are
counted before or after they have grown together. (Sources here
Mf and Kz.)
29 vertebra + 3 to 5 caudal vertebra ( here assumed 5)...... 34
29 bones in the head............................................................
29...Sum 63
1 breastbone + 24 ribs.........................................................
25......................88
2 x 2 bones in shoulder girdle (clavicle + scapula)..............
4
2 x 30 bones in arm + hand..................................................60...Sum
64
2 x 3 bones in pelvic girdle (hip, seat and pubic bone)..........6
2 x 29 bones in legs + feet....................................................58...Sum
64.....128
Total: 53 + 43 + 33 = 216 = 125
+ 64 + 27 = 33 x 23 = 27 x 8 = 216
An anthropocentric history:

Fig Sk-20-67-1x
An anthropocentric interpretation, yes. The anteater
pangolin has 49 caudal vertebra!
10. Mouth - Teeth:
 Fig
Sk-21-68-1 Fig
Sk-21-68-1
Both the mouth and the enamel of teeth originate in chordates from
ectoderm, from the animal 00-pole. The new mouth of Deuterostomia
is formed through invagination of ectoderm inwards the front part
of intestinal canal.
The environment is primary anticenter and 00-pole to the organism
as 0-pole, and the direction inwards (pole 4a) through mouth is
expressed in the eating, also a stepwise building-in of the
00-pole into the center as one principle in the dimension model.
The direction inwards is also defined as primary
polarizing, splitting force, which becomes expressed in the chewing
of food, a decomposing process.
In the history of evolution teeth originate from a carapace skin
such as in carapace sharks (Placoderms), which later were
reduced to placoid scales in present-day sharks. Thus, they were
structures at the surface that later immigrated into the mouth cavity.
The osteoblasts, the kind of single cells that build skeleton,
form also the hard material of teeth and originate as said above
from the neural wall in he embryo, i.e. from anticenter to the neural
tube at animal pole. (While the inner, sensitive dental pulp derives
from mesoderm.)
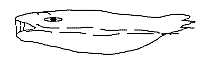
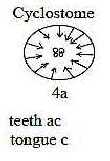
In an earlier species as cyclostomes, e.g. present-day lampreys,
the mouth lies in the surface plane with circles of horn teeth pointing
inwards the tongue in the center - with 1 tooth on the tongue! The
opposition in tongue muscles from mesoderm (originating from vegetative
pole) and teeth from around the animal pole is here very clearly
expressed in the center-anticenter polarity, in this case in the
2-dimensional plane.
 Fig
Sk-22-69-2a Fig
Sk-22-69-2a
In crustaceans one tooth immigrated as far as down into
the stomach - a polarization center - anticenter among teeth!
A precursor to the cyclostomes as the lancelet
without a cranium had a "wheel organ".
We can note that later evolution implies polarizations
from the circular structures into lower-upper jaws, ventral - dorsal
rows of teeth, in directions upwards - downwards etc.
The polarity in origin of tissue between tongue and teeth may be
seen expressed in their main, later functions:
- Teeth — Eating =
inward direction.
- Tongue — Speaking
= outward direction. (See Language, Speech
organs.)
Differentiation of teeth, dimensional forms:
They correspond to the form of contacts with food: points-linear-2-dimensional:
0 →1 →2:
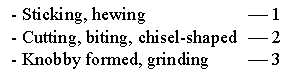 Fig
Sk-23 . Fig
Sk-23 .
In early chordates all teeth were approximately of the same kind.
They became more and more differentiated during the evolution towards
mammals. Bony fishes and reptiles had only teeth to grip and cut
with. Thus, the evolution should dimensionally have been of the
forms 3 ←2 ←
1 to the knobby molars.
About number 3:
In the early subclass of mammals, "Prototheria",
the molars had 3 nibs in a linear row. In later mammals, "Eutheria"
, these knobs formed a triangle, hence were 2-dimensionally arranged.
Apex of the triangle pointed inwards in upper
jaw, outwards in lower jaw (Fz p. 144), a detail that also
seem to express the fundamental opposition between directions in
the polarity dorsal - ventral and original 00- and 0-poles of the
embryo.
END
|