|
- Sense of pain is phylogenetically an old sense.
- Nerve fibers for pain exist in all kinds of membranes and
tubes (Mf), hence what may be regarded as the tissue
level underlying level of organs. (Embryos in their first
stages consist mostly of membranes and tubes before a nervous
system is developed.)
Pain is a general sense with little specificity, primarily
a chemical sense at destruction of cells but may be activated
by temperature and pressure as well.
The function of pain is to preserve the
organism as an integrated whole; raggedness and breaks gives
pain. It's a witness of the integrating force of an organism,
hence at bottom of all differentiations within its body. In
terms of the dimension model the sense of pain can be regarded
as an expression for the primary binding force from 0-pole
at polarization of 5th dimension degree (shortened d-degree).
On the chemical level destruction of a cell membrane leads
to outflow of K+-ions. There is a
strong connection between quantity of K+
in the intercellular tissue fluid outside the cell and the
intensity of pain (LEL p. 170); thus it's a chemical
expression for a destroyed membrane. It's mainly the level
of K+ inside a cell that decides
its rest potential.
At propagation of all nerve signals there
is an outflow of K+, however in very
small quantities, followed by an inflow of Na+.
It would be possible here to see a connection between nerve
signals as such and the sense of pain, where the counter directed,
opposite inflow of Na+ is lacking.
Pain becomes a one-way direction force outwards. (Cf. mental
pain when the "I" doesn't meet confirmation from
others.)
Another similar example that connects pain
with direction and position of otherwise usual substances
in the nervous system is Acetylcholine, a very old transmitter
in the history of evolution. It's found in a multitude of
synapses - inside nerve fibers and with a very short life
time outside in the synapses. Applied on the skin in a high
dose it gives pain. As in the case for K+
the condition is an unusually high concentration and
localization outside cells, i.e. outside the regulation in
nerves and synapses.
[There are obviously exceptions
from the rule that innervation for pain sensations exists
in all membranes and tubes. Evidently it's possible to cut
and burn in bowels without pain for the patient, while stretching
lengthwise gives pain. Could it have an embryological
and geometrical explanation? Pain from stress that can hurt
their original geometry? Intestines have their origin from
archenteron and primary vegetative 0-pole of the embryo, implying
the character of outward direction (divergence) in d-degree
4 (0 → when unnaturally reinforced giving pain? While
outer skin derives from the animal 00-pole, with circular
geometry in d-degree 3, when broken giving pain?]
Other features that point out pain as a fundamental sense
of high d-degree in our model:
- Pain is a sense with only free nerve ends, more or less
branched. The other general senses have free nerve ends too
but have also developed encapsulated ends of specific types.
It indicates a way from a primordial, more
elementary radial structure towards differentiations, in geometrical
terms of our model from vectors in d-degree 4 to the polarity
circular - radial in d-degree 3.
- Part of the nerve ends react to several kinds of
stimuli, both to chemical ones, temperature and pressure,
are "polymodal". Others react only to specified
stimuli as high temperature or strong pressure (Nf),
which indicates a step to differentiation, halfway to the
following senses - as a result of "polarizations".
- The nerve ends don't adapt, which seems natural with respect
to their function to preserve the organism. In relation to
adapting senses it's like a mathematical function relative
its derivative.
- Pain has the steepest log-curve of all the senses. (All
senses are logarithmic in the relation between intensity of
the stimulus and perceived intensity.) Log-curves of the senses
ordered after steepness:
Pain - Heat - Pressure - Cold - Vibration
- Hearing - Light (Nf).
Pathways of the pain-conducting nerves are special:
- Most curious - if shown to be a fact - is that the nerve
fibers for pain seem to enter also ventrally (Nf p. 461)
into vertebra (while all other sensory nerves enter only dorsally).
It looks like a reminiscence of the underlying two-way direction
of d-degree 4 in our model?
- Pathways for the general pain - and temperature - pass in
a special lateral tract in the spinal cord, nearer the ventral
horns, the paleospinal tract, phylogenetically older
than the dorsal tract in which nerves for other senses pass
as for lower pressure, touch, vibration and motions in joints
etc.
The nerves for pain are also more disordered.
- It should be underlined too that this paleospinal tract
- via the limbic system and thalamus - spreads out the signals
widely to the whole brain - as a radially directed vector
field.
C-A-delta fibers:
That the sense of pain include also a half step towards senses
"of lower degrees" is evident from the two kinds
of nerve fibers for pain: C and A, corresponding to a step
from a more general (diffuse) pain to a distinct:
- C-nerves, unmyelinated, propagate the "slow pain".
They pass as mentioned in the paleospinal tract and have the
general, divergent distribution.
- A-delta nerves for "fast pain" are myelinated
and pass through the neospinal tract to special areas
in primary sensory cortex with its map of the body.
C-threads only branches, A-threads form
plexa, more complicated structures (Nf p. 466).
C-threads lie also deeper in the skin than
the A-threads.
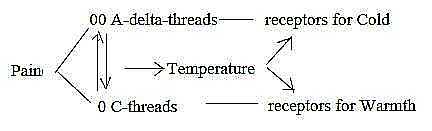
Pain - Temperature:
A step of polarization seems obvious between pain and temperature.
The sense of temperature too has C- and A-types of nerves
but here polarized to complementary receptors, with C-threads
for warmth and A-threads for cold (TA p. 73). Cf. that
strong warmth gives pain and that this sense had the next
steepest log-curve:
 Fig
Gs-1-127-1 Fig
Gs-1-127-1
This implies also a d-degree step as from a function to its
derivative:
- Pain receptors are tonic, not-adapting, always with
a certain activity.
- Thermal receptors are "phase receptors",
fast adapting, reacting to changes, the "derivative"
type (Aph).
Types of pain and muscle tonus:
- Superficial, shrill, burning, shooting, localized pain raises
blood pressure and muscle tonus, effects like those of the
sympathetic nervous system.
- Diffuse, dull pain from deeper tissues lowers muscle tonus
and blood pressure: effects like those of the parasympathetic
system (LEL).
Hence, it seems as the two reactions can
be described in terms of opposite directions:
- Pain from anticenter: 00 —> 0 : Sympathetic nervous
system as center-pole activated: —>0.
- Pain from center: 0 —> : Parasympathetic
nervous system as anticenter-pole activated: <—
00.
The inhibiting system is of the "antiparallel"
type as for the other skin senses, with signals coming from
higher centers, mostly centers in the brain stem near the
aqueduct between 4th and 3rd ventricles (LEL p.171).
Lateral inhibition doesn't exist, but activation
of other skin senses as touch may have some inhibiting effect.
2. Temperature:
Shapes of receptors and functions:
As mentioned above the sense of temperature
has free nerve ends as those for pain but also, according
to older sources (Zf ) specialized, encapsulated end
organs. This latter apprehension may have been revised but
mentions round capsules (Kruuse's) around branched
nerve fibers for cold and more oval or banana-shaped
capsules for warmth - around a more horizontally branched
nerve fiber (Zf p. 268).
Cold
Warmth
 Fig Gs-2-128-1
Fig Gs-2-128-1
Such a geometrical polarization in shapes resembles those
between sacs in the sense of equilibrium and the windows in
cochlea, the organ for hearing. It may be apprehended as expression
for an angle step outwards in d-degrees, as from 360°
towards 180°.
Cf. temperature as degree of spread in molecular
velocity: colder = decreasing spread, warmer = increasing
spread. Vertical axis = number of particles, a principal sketch:
 Fig
Gs-3-122-1 Fig
Gs-3-122-1
Higher warmth implies more of motions, of kinetic energy,
the direction towards increasing entropy. With this aspect
cold comes to represent the 0-pole, warmth a relative 00-pole,
~ lower d-degrees.
As mentioned about pain the receptors for
temperature are of the derivative type, reacting to changes
- and even to the the velocity of these changes (Zf).
It should be observed here that they react to the direction
of these changes: cold receptors on decreasing temperature
(~ convergence), receptors for warmth on increasing temperature
(~ divergence).
Cf. contraction of blood vessels as reaction
to cold, dilation of the vessels as reaction to
warmth. It confirms the view on the polarity in figure Gs-1
above.
- A-threads, myelinated: convergence from a 00-pole towards
cold.
- C-threads: divergence from a 0-pole towards warmth.
Myelination is also a later development.
(It has been shown however that C-threads
also can relay cold but then as it seems at much lower temperatures
than the A-threads (Nf p. 443). Could it be a reminiscence
of an older system with only unmyelinated C-threads and their
general spread in the brain?)
Receptors for cold lie deeper in dermis
(or subcutaneous layer (Zf p. 268) than the receptors
for warmth.
Another information (Nf) may be interesting with the
dimension chain in mind: The sensitivity measured as threshold
stimulus of receptors are 4 times higher for warmth than for
cold (0,001° relative to 0,004°), a relation 4/1
that can illustrate increasing differentiation towards lower
d-degrees.
Temperature intervals:
Receptors for cold answer in a temperature interval roughly
15° to 35° C, receptors for warmth roughly between
20° to 45° where they finish answering (Nf p. 442).
Hence an overlapping interval 20° - 35°. The latter
receptors have maxima around 38° to 43°.
However, cold receptors get activated above
45° together with specialized pain receptors, which shortly
can give what is called "paradoxical cold" sensations
from heat objects. In the central nervous system there are
also nerve cells that get impulses from both cold and
pain neurons that get identified as heat (Mf p.
313 f).
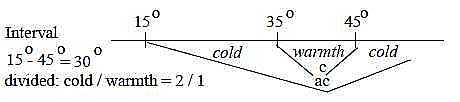
Fig Gs-4-129-1
One interpretation could be that the activation of cold
receptors occurs through pain as underlying level as shown
with dashed arrows in the figure below - in agreement with
the dimension model and with interpretation of pain as the
general sense polarized in the following ones.
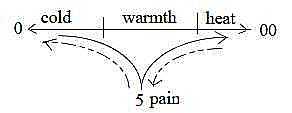 Fig
Gs-5-129-2 Fig
Gs-5-129-2
Or with a related view: The presumed "pole exchange"
in a d-degree step 1 → 0/00 (represented in each
step of a dimension chain) should imply that heat as divergence
defines a secondary 00'-pole, redefining cold on the thermal
scale: cold as direction inwards the 0-pole. (5th d-degree
equivalent with 0/00, d-degree of motions.)
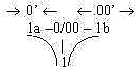 Fig
Gs-6 Fig
Gs-6
Receptors as a keyboard for temperatures:
The receptor threads registering temperature are differentiated
with their maximal burst frequencies at different degrees
of temperature (Nf p. 442), reminding of hair cells
in response to sound waves. How this differentiation is organized
in cochlea is well understood, but how among receptors for
temperature? (No information in used references.) Perhaps
through a map of positions within different domains in the
skin or inner membranes - with a corresponding one in the
brain?
3. Pressure, Touch, Vibration - mechanical senses:
The senses for pressure, touch and vibration are mechanical
and appear as a further differentiation from the sense of
pain. Cf. the differentiation of pain above as a) polymodal,
b) for warmth, c) for pressure, and the order of steepness
in log-curves: pain →> warmth →> pressure.
- Nerve fibers for pressure are mostly thicker, myelinated
threads in opposition to the thin threads for pain and temperature
(Mf).
- The pathways for most of these mechanical senses go in another,
more distal neospinal tract in the spinal cord than
the more lateral, ventral tract for pain and temperature,
the paleospinal tract, which also is the one for hard
pressure (Aph).
Ends of nerve fibers:
Free nerve ends exist as for instance around roots of the
hairs, which are encircled by nerve ends. Mostly however,
the nerve ends for mechanical senses are encapsulated in capsules
of connective tissue and further differentiated in specialized
structures in at least 4 known types.
We can identify 2 polarization steps:
- in slow and fast adapting ones (tonic and phase types),
each of which polarized
- in those with small and those with bigger receptive fields.
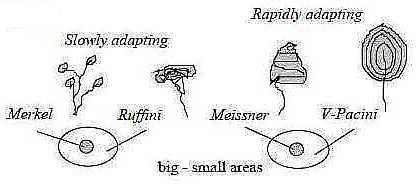
Fig Gs-7-130
Further, there is the differentiation in function between
fine touch and pressure (only a difference of degree) and
deep pressure.
The fast adapting end organs are simultaneously
sensitive to vibration and differentiated between low and
high frequencies (f.) of vibration.
1. Slow adapting, simpler kind of embedding:
a) Merkel's discs : Receptive fields small (~
center pole)
- Structure: Dendrites disc-shaped, closely ("vertically")
attached to special big cells in epithelium.
- Sensitive to fine touch and pressure.
b) Ruffini's corpuscles: Receptive fields bigger,
vaguely demarcated (~ anticenter pole).
- Structure: Nerve ends spread in a bundle of "horizontal"
collagen fibers.
- Sensitive to pressure and distortion, tension of skin. Note
this angle step.
2. Fast adapting, more complex kind of embedding:
a) Meissner's corpuscles: Receptive fields small
(~ center pole).
- Structure: doubly embedded, branched and coiled dendrites
with ends surrounded by modified Schwann cells and this whole
enclosed in a capsule.
- Sensitive to fine touch and low frequency vibration.
b) V. Pacini's corpuscles: Receptive fields
big (~ anticenter pole), vaguely demarcated.
- Structure: one single dendrite thread within several layers
of concentric collagen fibers (lamellae), rather flat.
- Sensitive to deep pressure and high frequency vibration
(~ "overtones").
It's noteworthy that the structures differ more clearly than
their functions seem to do.
If we should try to apply the dimension
chain on these separate structures, certain features at least
are possible to identify as such:
- The relation between "radially" spread discs of
Merkel's type versus the horizontally arrangement of Ruffini's
type as poles from step 4 →> 3.
- The big rounded, doubly embedded Meissner's type versus
the more flat concentric Pacini's type as a feature relation
3 to 2 (and 2 to 1 with regard to the structure of dendrites
within the corpuscles).
It has been proved that it is the embedding
in corpuscles of the nerve ends that makes them fast adapting.
The fast adapting types, sensitive for vibration, could perhaps
also be described as derivations from the slow adapting in
two steps (?):
4 -|- 3: Merkel's: radially branched
nerve ends: fine touch and pressure.
↓→ Meissner's:
branched, coiled nerve ends: fine touch and low f.
vibration.
3 -|- 2 - 1 Ruffini's: horizontally nerve
end(s) in bundles: pressure and tension.
↓→ Pacini's:
one single, vertical nerve end: deep pressure and high f.
vibration.
Positions in layers of skin:
Pain: - Merkel's — Ruffini's + Meissner's
- Pacini's: →> more embedded:
If we may regard the senses from pain to the mechanical senses
touch and pressure as differentiations analogous to steps
towards lower d-degrees, from free nerve ends to big end receptors
and from non-adapting to fast adapting, the direction in positions
becomes roughly from outer layer of the skin to inner, underlying
layers. Epidermis originates from the animal 00-pole, deeper
layer from endoderm of the vegetative 0-pole and mesoderm
embryologically.
The localization of receptors in opposition
to the suggested d-degree of their character could perhaps
be understood in terms of lacking answers:
- Pain as outward directed, not answered
by adequate response from outside, outer skin.
- Deep pressure as inward directed, not
answered by balancing pressure from inside.
4. About receptors in joints, an addition:
Besides other sensory receptors within the body as e.g. baroreceptors
in blood vessels and proprioceptors in muscles and joints
etc. there are receptors similar to Pacini's and Ruffini's
in joints (Nf p. 440). Those which adapt slowly give
information about direction of motion, velocity and position.
- They react to motion in the joint within a certain angle
interval, a differentiation of receptors as along angle steps
in a dimension chain: change of Direction, ~ d-degree 4.
- Frequency of receptive response is linearly proportional
to the velocity of the motion. Velocity has in this dimension
model been suggested as the physical quantity of a dimension
step, type 1→>0/00 (the very quantum jump between
d-degrees.
- Discharge of the receptive response drops to a level that
corresponds to the position of the joint, hence a spatial
qualifier as out of angled directions; position, d-degree
3.
The same receptor mediates information of
all three properties: direction, velocity and position:
D-degrees in the dimension chain:
4 —————————————3
Direction Velocity Position
5. All senses: dimensional aspects on their mutual relations?
Usually in human biology there is talk about 5 to 6 senses
(at least the hitherto identified):
Equilibrium - Taste - Smell - Sight - Hearing
plus
Kinetic sense from inner muscles and joints.
To these comes the skin senses for pain - temperature - touch/pressure
- vibration, plus all inner receptors for blood pressure,
chemical milieu etc.
An interpretation of all these senses in
their mutual relation with aspects from a dimension chain
becomes probably most natural with a division in kinds of
stimuli they respond to, and how these stimuli are related
to fundamental physical qualities (or "quantities").
(About how these physical qualities are suggested to be stepwise
defined in the dimension chain, see here.)
Sense of equilibrium is connected with gravitation
and outward acceleration, chemical senses with mass and matter
in next steps (and charge), sight with electromagnetic waves
related to charge etc.:
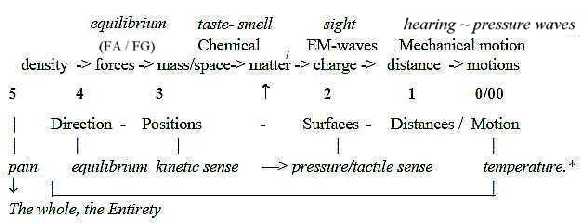
Fig Gs-8-132-1
* D-degree step 1 →> 0/00 debranched from step
5 →> 4: Temperature as warmth is related to density
among particles, partly a matter of their motional energy
and velocity (a quality distance/time), partly a matter
of imbalances in radiation (EM-waves).
Step 1→>0/00 debranched from
step 4 →> 3: motions of the body registered by the sense
of equilibrium and receptors of the kinetic senses.
With the loop version of the dimension model
the senses regarded in such a chain becomes also roughly an
illustration of steps from inner senses to outer "near
senses" as taste and smell to outer "distant senses"
as sight and hearing:
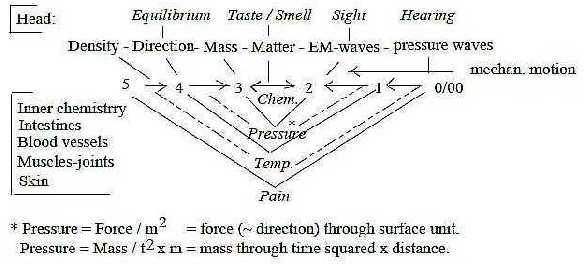
Fig Gs-9-132-2
Thus, pressure may be translated into a relation force (4)
and distance (1), or to mass (3) and the quantities distance
plus time.
Directions and sense of Equilibrium
are expressed in motions of the body and closely related to
Sight.
Mass →> Matter as Chemical senses:
body motions governed by Smell and Sight in seeking for food.
|