|
Most data here concerns the human eye.
1. Geometry of analysis:
A geometrical analysis of received pictures is performed by the
eye. It appears to be much of the same kind as the elementary geometrical
definitions of the complementary poles in our dimension
model.
Scientists have been able to distinguish 5 different
types of ganglion cells, sensible for different geometrical features
(Nf p. 373), in number also the same as degrees in the model.
There are for instance cells sensible for direction
of linear structures, others sensible for round forms, others for
curves, convex or concave, others for straight lines in different
angles, others for motions of geometrical elements in certain directions.
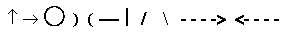 Fig
S-1-113-2 Fig
S-1-113-2
Cf. in the model proposed geometries of polarized dimension degrees
(shortened here d-degrees) d-degrees:
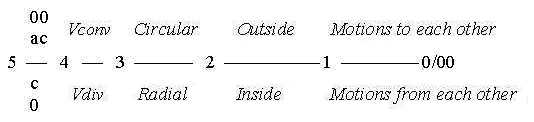
Fig
S-2
Direction (4): Vdiv - Vconv, Volumes (3): radial
- circular structure, Surfaces (2): concave - convex, inside
- outside. Distances (1): motions to - from each other (0/00).
2. Number of cell layers:
There are 5 layers of cells in the retina (compare number
5 in the many other cases of arrangements in the human body):
3 cells in 'vertical' direction, from receptors
of photons, the cones and rods, via bipolar cells to ganglion cells,
leading the signals to the optic nerve and visual cortex in the
brain.
2 intermediate transverse layers of cells, at
right angle to the other ones, the layer of horizontal cells
and the layer of amacrine cells.
It's a number relation as in step 3-2 in the dimension
chain, where the polarization 3 →
2 was defined in elementary geometry: radial versus circular, outer
poles of d-degree 2 in the model, that of surfaces.
An outline of the layers without any precision:
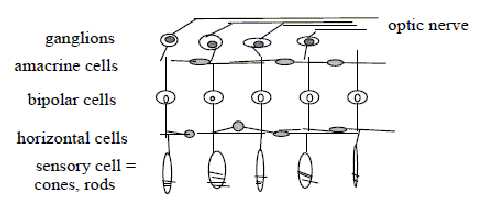
Fig
S-3-111-1
Here the cells in 'vertical' lines are the integrating ones. Cf.
the relation between d-degrees 3 and 2 as corresponding to a 0-00-relation:
the 0-pole as integrating, the 00-pole as differentiating. Bipolar
cells gather signals from many receptors, ganglion cells from many
bipolar ones.
Cells in the transverse, 'horizontal' layers have
a similar function of lateral inhibition or stimulation as corresponding
cells in the general nervous system: facilitate or inhibit the sensitivity
of propagated light signals: it's the polarization we have suggested
as d-degree step 2-1 in the chain of polarities
in the nervous system.
|
5 —>
4 —>
3
\
1 \
2 → debranched
d-degrees
meeting the other way around
|
Fig
S-4 |
3. Center - anticenter polarity in d-degree 2:
Structure of the eye is on a macro-scale 3-2-dimensional with the
globe (3) and the retina (2): pictures principally 2-dimensional.
We could note that the eye-globe really can rotate, the "external
motion" assumed in d-degree 3 in our model (!). A rotation
through its 6 muscles (m.), a number that happens to be sum of the
poles 3a + 3b out of polarized d-degree 3.

Fig
S-5 (after Mf p. 335)
(Cf. muscles in the dimension chain
of organs identified in d-degree step 3 - 2 and angle steps
180°→>90° →>
45°.)
A general, structural feature on a more detailed level regard surfaces
(d-degree 2) and the polarity between a central area and an anticentric
one; not explicitly included before in the model among the polarizations
of d-degree 2 (concave / convex, inside
/ inside...).
 Fig
S-6-111-2 Fig
S-6-111-2
The same polarity appeared among types of corpuscles for general
senses in the skin but only as smaller and bigger fields
for reaction. In the eyes this opposition becomes much more obviously
a polarity. (Embryologically
the differentiation of cells around animal pole in the blastula
is of the same kind.)
- The pupil of the eyeball is of course a first example
of this center - anticenter (c - ac) structure. The center as just
an opening, a hole (!) could be regarded as one kind if inversion,
cf. the cell
interpreted as inversion of an atom.
Area of this center, the pupil, is governed by
radial and circular muscles, the geometric poles 3b and 3a out of
d-degree 3 and outer poles of d-degree 2 in the dimension chain.
These muscles are in their turn governed by the
sympathetic
and parasympathetic nervous system respectively for outward
and inward directions, The fact that the pupil not only functions
as an aperture but also chronically "oscillates" a little
in size (Nf p. 353) could perhaps be traced back to the vibrational
moment assumed in d-degree 4 of Direction, origin of these muscles?
- Next example is the organization of cones and rods in
the retina, with cones in the central region and fovea, a central
shallow depression, and rods in the anticentric area. While cones
have more own pathways, signals from up to 1000 of rods (Aph)
may converge to one ganglion cell: it shows the typical convergence
= direction inwards from an anticenter. The whole system for analyses
of signals seems built on processing steps of convergence /
divergence (Mf p. 363), the forces from 00- and 0-poles.
1/3 of the fibers
in the optic nerve comes from the fovea, 2/3
from the surrounding area (also a noteworthy division) as numbers
of steps from d-degree 3 (?).
About P and M in the figure, see below.
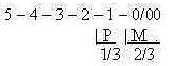 Fig
S-7- 112-1 Fig
S-7- 112-1
In cortex however, it appears that these threads occupy equal parts
of the primary visual cortex (Mf p. 310), a division 1/1
which could support the view on a c-ac-polarization of the same
d-degree.
We could note the number 3 too in the different
kinds of cones creating all colors.
White light is all wavelengths gathered together: the eye shows
the generalized peripherally in the sensitivity of rods, the separated
wavelength, the colors, centrally, which reflects the polarity unity
- manifold between 0- and 00-poles.
- The "P-M-system" is a third example of the c-ac
polarity in d-degree 2. The reception areas of the ganglion cells
are usually circular with a center field and surrounding anticenter
field:
Ganglions for cones are called P-cells: they have
small reception areas, mostly in the center.
Ganglion cells for rods, called M-cells, have
larger reception fields, mostly peripherally.
That this division corresponds to a polarization
becomes evident too from the fact that they have synapses in different
cell layers in the visual cortex and get projected to different
secondary areas (TA p. 106).
These types show simultaneously a relation of higher d-degree to
lower ones in two respects, as such a relation always is of the
kind 0 to 00:
The P-cells react especially on colors,a property
connected with surfaces (~ d-degree 2) and wavelengths (~ d-degree
1 as distance), the M-cells especially on motions, the last degree
0/00 in the dimension chain; cf. the figure
above.
Further, M-cells adapt fast in opposition to the
P-cells. Such fast adapting cells can be regarded as "derivative
cells", illustrating changes in the same way as the derivative
of a mathematical function illustrates changes of directions in
a curve and implies a decrease in d-degree.
4. On-off-center cells:
The polarization of the derivative M-cells show the complementary
feature of poles in our model very clearly.
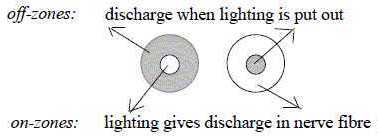
The receptor fields of the ganglion cell becomes
through different connections in the net polarized in two kinds,
on- and off-center cells:
One type gets active when light hits the center,
and inhibited when it hits the circumference of its receptive field.
The other type is the inverse: gets active when
light hits the surrounding area, inhibited when it hits the center
of its receptive field.
Fig
S-8-112-3 after Nf p. 370
We may compare with polarization of a derivative in
an inclination upwards of a curve and an inclination downwards,
with polarization of signs plus/minus
as in the physical property of charges.
According to a principal sketch (Nf) inhibiting
pathways go to centers in the one kind, to the anticenter in the
other kind.
The arrangement could be interpreted in terms of the final step
1 →> 0/00
with "pole exchange" in the dimension model: Pole 1b,
"motions from each other" out of divergence and 0-pole,
define an anticenter, a new 00-pole, and "motions to each other",
out of convergence and 00-pole, define a new center: With 00-pole
representing inhibition, 0-pole excitation we get following scheme:
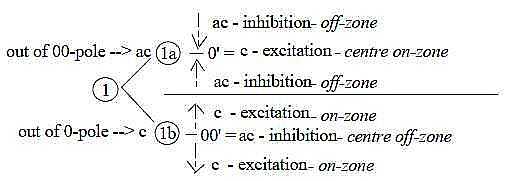
Fig
S-9-113-1
The approximate number of cells reacting in different
ways seems to include 3 steps of divisions in percentage:: 100 →>
50-50, 50 →> 30-20, 20→>(10-10 ?, no figures given):
- 50 % of them react both when light is turned on and when it's
turned off, on-off-cells,
- 30 % react when light is turned off, off-center cells,
- 20 % react when light is turned on and during lighting, on-center
cells.
These 20 % are further divided in such that only react initially,
and those that also react during continuous lighting.
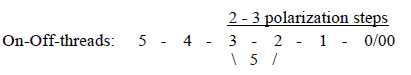
Fig
S-10-112-2
Since vision is built on light, on electromagnetic
waves and the electromagnetic force (FEM),
it is a reasonable question if the polar arrangements in the retina
in some way could reflect the continuous changes between electric
and magnetic factors in EM-waves. (In the interpretation of forces
we have assumed the EM-force defined in d-degree step 3 - 2 with
E and M as the complementary poles or fields.) One association goes
to the light and dark rings on a detector screen when photons pass
a small hole in a screen in experiments showing the double nature
of light.
5. Several inverse features appear in the sense of vision:
- The nerve cells get hyperpolarized (from ~ 40 mV to ~ 70 mV)
when excited - instead of depolarized as in other nerve cells.
- The photoreceptors secrete transmitters in darkness. (A condition
for the sight signals is a continuous own activity in the cells.)
- The receptor cells with their extensions are turned inwards
in the retina of mammals.
Besides that the pictures the eye delivers to
the brain are upside-down.
Such properties could eventually be connected with the turn in
main directions in the loop version of the dimension chain in step
3 - 2.
 Fig
S-11 Fig
S-11
(Or with "the pole exchange" in d-degree 0/00
of our model where pole 1b from 0-pole defines a new anticenter
00' and pole 1a from 00-pole defines a new center 0'. See file General
senses, figure Gs-6.)
6. Direction and distance to viewed objects:
These to properties in the visual field are registered in different
columns in primary visual cortex.
Direction seems defined through a difference
in the storage of cells that receive signals from the two eyes:
those from opposite eye lie gathered or tightly over one another
in the column, those from the same eye are more horizontally spread
(Nf p. 385).
In other terms, the opposite eye seems to represent
a center or the vertical axis, the eye from the same side the horizontal
one. It corresponds to first polarization in d-degree step 4 →>
3 in our model. The differentiation becomes horizontal for directions
as along a circle..
Distance in the depth columns seems defined
through equal, horizontal distribution of signals from both eyes
(a bit vague expressed in the reference). Differentiation hence
vertical. (It should also represent a step from vector to scalar.)
An own interpretation of the reference below:
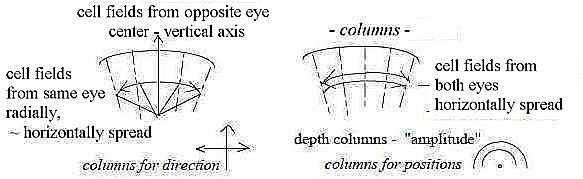
Fig S-12-114-1
Texts in this figure concerns actually cells in one single column.
It's said in more general words that the vertical organization in
a column concerns details and orientation, the topographic picture
of retina the "place"; cf. in this sense directions (d-degree
4) - of lines and other details too - versus 3-dimensional space.
7. Layers in columns of cortex;
The deeper the research towards a detailed level, the more a stringent
geometrical organization within biology is revealed. There is for
instance also a strict order of fibers in the optic nerve. At first
station where fibers cross over from one eye to the other side (lateral
geniculate nucleus) the cells are arranged in 6 layers. Fibers
from the opposite side go to layer 6 and 4 + 1, fibers from the
same side to layers 5 and 3 + 2. (Nf p. 375).
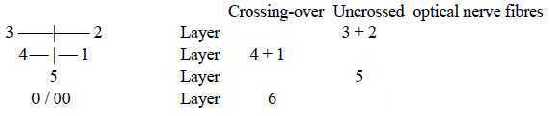
Numbers from reference.
Fig S-13-114-2
Cf. 3 polarizations of 5 in a dimension chain, 0/00
(~ "6th" d-degree) as one of them.
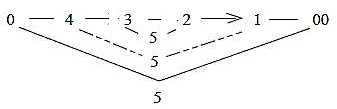
Fig
S-14-7-3
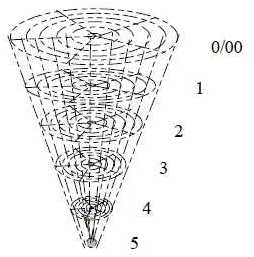
8. About number of cells in the human retina:
Human beings have about 125 million rods and about 6 million cones.
(Also about 6 million bipolar cells) and about 1 million ganglion
cells and fibers in the optic nerve (Aph). An interpretation
of the very numbers:
|
Levels as in a dimension chain = 6.
5 radii on each level.
Gives 125 crossing points.
+ 6 along the central axis = 131
Suppose each level represents a 10-power.
We get the number 125 x 106 rods
and 6 x 106 cones.
|
Fig
S-15-114-3
|
|