|
1. A fundamental invention:
Centrioles are among the most beautiful and clear expressions for
geometrical design within biology; an "invention" of eukaryotic
cells on the level of organelles.
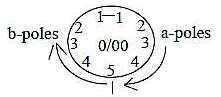
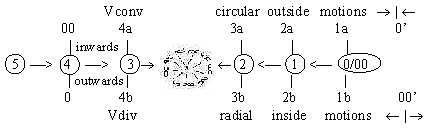
In cross-section the structure illustrates the center anticenter
polarity (0/00) in arrangement
of microtubuli and radial fields illustrated by the spokes, in our
model defining dimension degree (d-degree) 4 with outer poles 0
and 00, which meet in the d-degree of motions.
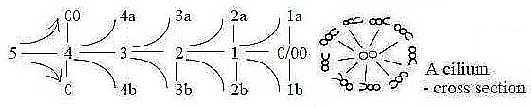
Fig Cil-1-157-1
Why the number 9 in the ring of tubules? Scientists dont seem
to ask such questions. For some speculative proposals, see the end
of this file.
Cilia and eukaryotic flagella, built on this plan, appear as first
"extremities" of cells. They serve a double function of
sensory antennae and external locomotion in the environment. Nearly
all mammalian cells have one cilium.
The inner centrosomes, consisting of two centrioles,
have been called the "heart" of the cytoskeleton system
in a cell, radiating through its cytoplasm.
Notice in the dimension model that d-degree 1 are debranched
in each step outwards and may polarize into the "d-degree of
motions".
 Fig
Cil-2 Fig
Cil-2
2. Evolution:
Evolutionary, such cell extremities have simpler forms in Archae
and prokaryotic cells, and the evolution may be described as steps
from center to anticenter or just to more complex c/ac-structures:
- from simple bundles of protofilaments in Archae (resembling
pili), as an original center, to
- central but hollow, bigger, screw-shaped tubules (center widened
to a circle) in prokaryotes, to
- the c-ac structure with explicitly anticentric manifold of
tubules in eukaryotic cilia, including spokes.
This evolution has a character of d-degree steps from center to
anticenter and simultaneously of geometrical steps of substantiation
3 ← 2 ←
1 in plain projection.
The differences include 2 polarities:
a) between Archae and the other: In Archae bacteria the microtubules
are build from below, i.e., from inside the cell outwards
(as a form of secretion). In pro- and eukaryotic cells they are
built from the top furthest out. It reveals a pole exchange
0 00.
b) Next pole exchange of type 0-00 appears between prokaryotic
and eukaryotic cells in the transportation of building material:
- in prokaryotic cells it occurs inside the hollow tubule,
- in eukaryotic cells outside the walls of the many
microtubules;
a polarization c-ac besides the one between one central tubule
and many smaller at anticenter.
Such features support a view in the development from center to
anticenter, principally outwards, during increasing complexity,
while simultaneously the direction in the dimension chain is inwards.
 Fig
Cil-3 Fig
Cil-3

Fig Cil-4
(The fact that these organs are not homologous but
built in different ways with different proteins, can support the
presumption here that its geometrical principles that reign.)
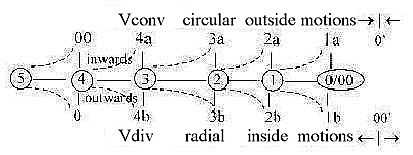
3. Polarities in eukaryotic cilia:
Transportation:
In eukaryotic cilia there is further the polarity between outer
and inner "dynein arms" of microtubules at anticenter:
- transportation upwards of building material occurs as said above
on inner side of microtubules,
- transportation downwards occurs on outer side of them, which corresponds
to the fundamental directions of d-degree 4 in the dimension model
(inwards from anticenter, outwards from center).
The outer and inner arms differ too in protein
structures (KI-O), hence in some respect obviously polar.
One reference mentions the proteins kinesin and dynein
as such mutual polar proteins in directions of transportation.
The polarity resembles the arrangement of vessels in plants:
outer phloem for downward transport and inner xylem
for upward transport. There is actually a similarity too between
the different structures of cilia in Archae, prokaryotic and eukaryotic
cells and the development of steles (a, c, e in file Plants,
No. 4). Plants as cilia of the Earth!
There is further the similarity that roots of trees
lack pith, and basal bodies of cilia "lack" the
central tubules.
Structural differences of anticenter:
The differences between centrioles in cilia, basal bodies and centrosomes
may be regarded in terms of d-degree steps:
a) Inside the cell, in centrosomes and in basal bodies
of cilia, the centrioles consist of 9 triplets, 9 x 3 tubules
at anticenter. Outside the cell surface, in cilia, they consist
of 9 doublets, 9 x 2 tubules The numbers 3 and 2 happen to
correspond to outer poles in d-degree 2 and 1 in the model.
Geometrically the difference corresponds to the
step from membrane surface (d-degree 2) to cilia as d-degree 1 on
a relative macro-scale.
b) The centrosomes (9 x 3 tubules) in the inner of the cell
consist of 2 centrioles (mother- and daughter cell) in a curious
90° angle to one another, which geometrically implies definition
of a 3-dimensional volume.
When not taking part in cell division one of these
centrioles migrates to the surface and transform to a basal body,
thus illustrating a halving as a polarization step from a
volume to a surface (d-degree 3 →
2).
c) The centrosomes as MTOC organelles, near nucleus, organize the
radial protein skeleton of the cell. Their function as vector fields
(d-degree 4a-4b as outer poles of d-degree 3) becomes obvious.
Eukaryotes appear as a new level in d-degree step 3 - 2 through
meeting of directions according to the loop model. Its possible
to imagine centrioles in this step as a complex center (0/00),
expressing the radial / circular
polarity among proteins in this step:
0 → →
00'/0' ←
← 00, a haploid chain,
which get a complex center in the middle, c2
 Fig
Cil-5 Fig
Cil-5
- In d-degree 3 it would give the triplets, function of center
(MTOC) for radial outward vectors, the cytoskeleton and the spindles
as vector fields at cell division.
- In d-degree 2 outwards it would give the doublets, function of
basal bodies and further the structure of cilia.
Concerning forces, the electromagnetic force (FEM)
has been assumed defined in step 3 2 in the dimension chain of
forces.
The radial cytoskeleton arranged by centrosomes have a direction
outwards from minus-charge at center, plus-charge towards cell membrane
as is assumed about charges from 0- and 00-pole respectively.
Centers and spokes:
Centers of centriole structures are of different kinds: in the
inner centrosomes as in basal bodies there are no visible tubules
but "something", not possible hitherto to identify. Hence
called "9x3 + 0" structures.
Cilia outside the cell surface are of two kinds:
the primary, non motile one, without identifiable center structure,
and motile cilia with 2 single tubules in the center. How explain
this difference?
One guess is that the "something" in the center of
inner basal bodies and centrosomes as 0-pole in the d-degree 4 of
vectors are less substantiated, not yet have got the ring-structured
mass through inward direction from environment as anticenter. Cf.
also number 9 and 11, section No. below.
Scientists would probably explain it as a question
of need for stability in outer motile cilia or something like that?
The function of these centers is not yet understood.
Spokes in the cross-section depart from tubules at anticenter
and end in "heads" (some thicker structures) about halfway
to the center. These spokes are unexplained too. Its difficult
not to presume some invisible spokes as vectors from the center
too, and the picture can remind of the FG-
and FA-forces in macrocosm: gravitation inwards
to mass of galaxies and outward acceleration force (FA)
of "invisible" Space.
4. Sensory and motile cilia:
The basic structure appears either as sensory antenna,
called primary cilia, "9x2 + 0" structures, or as
organelles for locomotion,"9x2 + 2" structures, the motile
cilia.
The types represent the fundamental opposition
of the nervous system in multicellular organisms. (All developed
neurons have one cilium and fibers for cilia are found on the neural
plate.)
Primary, sensory cilia:
As the nervous system guides inner processes on a multicellular
level, so seems primary cilia of cells in vertebrates and mammals
guide a lot of internal structural relations, positions and processes
in the body and coordinate a big number of signaling pathways (Wikipedia).
Failures in their structure seems for instance to have a role in
the position of heart to the right instead of left side in the body!
Maybe these regulating functions are just expressions for cilia
as structures of d-degree 1 on the cell level and the very jumps
or steps of polarizations in a dimension chain behind relations
in the organism?
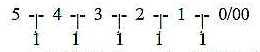 Fig
Cil-6-157-2 Fig
Cil-6-157-2
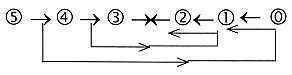
Each step:
 Fig
Cil-7-161-1 Fig
Cil-7-161-1
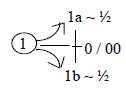
Cf. that d-degree 0/00 (or 00
in a haploid chain) as anticenter represents first polarizing force
in our model, and passive motions of these cilia are induced by
environment. Its also possible to think of poles 1a and 1b of d-degree
0/00 of motions in terms of insubstantial
"field lines" in the environment.
The primary cilia are used in more or less specialized shapes in
the primary senses of mammals, in photoreceptors in the eye, as
kinocilia on hair cells in the ear and on olfactory neurons in the
nose.
Cilia as primary receptors of sunlight in the eye
have a certain similarity with chlorophyll on the molecular level
in its circular structure, corresponding to the rotational construction
of porphyrins to anticenter rings around a center (a Mg-ion in chlorophyll).
Motile cilia and motions:
Centrosomes have been called motional centers inside the cells,
organizing the cytoskeleton. The motile cilia as first extremities
serve external locomotion, d-degree 0/00
in our model.
Developed cilia contain over 600 different proteins (Wikipedia).
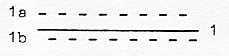
However, it has been shown that 2 proteins is enough to bring
about the beating motions of flagella (= cilia in eukaryotes). The
2 proteins could be illustrated by the poles 1b and 1a in the figure
above. Its added that many of the over 600 individual proteins
can function as small "nanomachines".
Flagellum of human sperms is actually a modified cilium.
(Cf. the mass number of side chain of amino acid Arginine = 101,
said to be richly found in fish sperms. About number 101, cf. the
ES-chain in the
genetic code.)
The motion starts at the very tip of cilia furthest out
according to observations. Curiously enough, since the "motor"
is positioned at the base of cilia. However, in our model motions
are the very end of a dimension chain outwards.
Tubulin makes up the building blocks of microtubules. It's
a globular type of protein (like F-actin in muscles), which
can illustrates quantification of a line in "half steps"
into motions..
 Fig
Cil-8 Fig
Cil-8
The polarization into "half steps": the poles 1a, "motions
to each other", and 1b, "motions from each other"
in the model, can give the pendulation as between two poles of convergence
and divergence. The figure could illustrate motions along the tubules.
The cilia describe externally a 2- or 3-dimensional
motion: a planar, wavy one or in other cases a more 3-dimensional
one with power and recovery strokes (Wikipedia). These d-degrees
of motions are in the model associated with d-degrees 3 and 2 respectively
(cf. No. 3 above). Which factors that decide the difference of motional
degree seems still not clear.
Several internal motions in the tubular structures
are described: a sliding or clipping force at positions of ATP,
also an oscillating movement along the whole length (KI-O).
Further, a rotating or twisting force (KI-O).
The microtubules have an ability of contraction too (Zf).
Its the two dynein arms in eukaryotic flagella that get the microtubules
in the doublets to slide against each other, which gives the external
motion (Wikipedia).
Thus, with all these sliding rotating or twisting oscillating
- contracting and clipping motions it seems as if they
could represent all the presumed motional moments in different d-degrees
of the dimension chain:
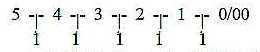 Fig
Cil-9-157-2 Fig
Cil-9-157-2
(D-degrees of motions in the model:
Vibration 1, rotation 2, spiraling 3, pumping as expansion/contraction
4, and "pole exchange" as first embryonic germ to motion
in d-degree 5.)
About details, there are at least 9 different types of dynein proteins
(KI-D, KI-O), eventually differentiating the microtubuli
in degrees of stiffness and sliding velocity as it sounds . Connected
with different d-degree steps?
5. Number 9:
Why this number 9 (x 3, x 2) of peripheral microtubules in centrioles
and cilia structure? As said above, scientists dont seem to ask
such questions. A random number?
Its a general hypothesis in this work however
that such numbers can have a deeper foundation. We may repeat that
this number has nothing to do with numbers of chromosomes or number
of protein threads in cytoskeleton arranged by centrosomes.
Naturally, following different suggestions are
only speculations.
a) Reading each step in the dimension chain as 2-figure
numbers, the difference outwards inwards in directions = 9:
5 | 4 | 3 | 2 | 1 | 0/00
54-45, 43-34, 32-23
10-01 = 9.
It would agree with the interpretation of motions in d-degree 0/00
as turns in main directions.
b) Compare number 11 = 54-43, 4332
21-10: the difference
between steps in outward or inward direction, i.e. in one-way
direction.
Note that 11 is the number of protofilaments in
almost all flagella of prokaryotes (Wikipedia).
(If number-base systems (nb-x) should be connected with different
d-degrees — cf. the genetic code, part III — 9 in nb-10
= 11 in nb-8. A d-degree step between pro- and eukaryotic flagella
types - ? - simultaneously with a step from fiber level to first
tube form, d-degree step 1 to 2 ?).
c) Number 2 in center of motile cilia: at each d-degree
the difference in directions inwards higher d-degree and lower one
is 2, also marking a difference of directions: 5 ←
4 | 4 → 3, read as 45-43
etc. A factor of opposite directed motions in each "quantum
jump" between d-degrees?
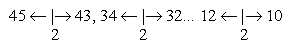 Fig
Cil-10 Fig
Cil-10
d) Number 9 as inversion of a series of 10-power displaced
11111
?
9 /\ 0, 1111111
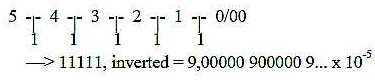
(5 ten-powers as steps.)
Fig Cil-11-157-2
e) Doublets as 99?
101 /\ 99009900
Cf. about side chain of Arginine above = 101 and last number
in the ES-chain, the genetic code..
f) Five steps of 1 (degree) squared:
| 111112 = 123454321
= 9 figures: |
|
Fig Cil-12-157-1b
9 figures from 1 to 5 inwards and back outwards. First and last
"1" in the number as poles 1a and 1b in the dimension
chain. This interpretation should imply that the total circumference
of microtubules had opposite poles and identifiable half turns in
some way. (Cf. the curious angled copying of the daughter cell in
centrosomes?)
g) The d-orbitals for 10 electrons in the periodic
system is the middle one in the 2x2-series
(x = 5 - 0) behind this system. There is 1 central ring-shaped electron
orbital and 9 electrons divided, 3 on coordinate axes, 6 on pairs
of plane quadrants. Thats 9 polarized, more peripheral orbitals.
Could centrioles as developed in the middle step according to
the loop version of our model have a deep correspondence in this
d-orbital?
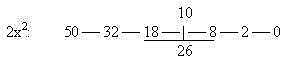 Fig
Cil-13 Fig
Cil-13
Its said that the doublet of microtubules in cilia consist of
13 threads in the A-rings, 10 in the B-rings (http://www.solunetti.fi/se/solubiologia/varekarvat_1/2/Cilier):
1/2 x 26 =
13 for A-rings with dynein arms and bridge to B-ring of next doublet
with 10 threads?
(9 x 23 = 207 = number for A-base
in nb-8; an association to the genetic code.)
h) 9 triplets in the volume of cytoplasm = 27 microtubules
= 33
9 doublets = 18, outside the cell membrane,
d-degree 2 = 2 x 32
A step 33 ← → 2 x 32 (18 in 2x2-chain
above)
j) Simple reading of numbers in last d-degree step of
the dimension chain:
Step 1 → 0 as 10: 10
-/+1 (as 1a 1b?) = 9 and 11, difference
2?
1b-poles define a new anticenter, ~ the 9 doublets.
1a-poles define a new center, ~ + 2 central tubules ?
Whatever to believe about the numbers, centrioles and cilia structure
seems to be the way of atoms to tell us a fundamental story about
life.
|