|
1. Levels:
In the dimension model it's assumed that each step in a fundamental
chain can develop to secondary whole dimension chains and so on,
thus to "level chains".
- Levels in accordance with a dimensional chain - a "level
chain":
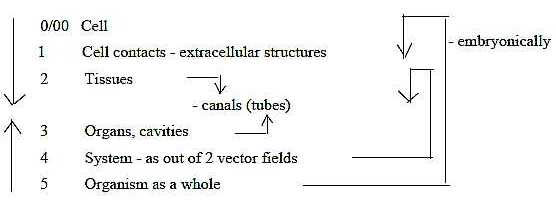
-
-
In physical and geometrical quantities:
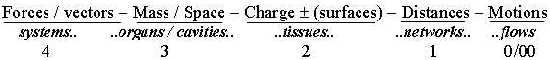
-
Fig L-2-29-2
-
-
Poles:
- c-ac: organism - environment,
- 4b/4a: nutrition system <—>
nervous system, inward/outward directions
in the two systems,
- 3b/3a: mass - space (~ cavities) in
the design of organs and tubes of the body,
- 2b/2a: differentiation of tissue layers
in e.g. inner/outer layers ,
- 1b/1a: negative /
positive affinities between cells, one expression for "movements
to/from each other ". or
2a - 2b: differentiation of charge over tissue layers and e.g. outside
- inside
1a - 1b: convergent flows and divergent along transportation nets.
Steps in a dimension chain in direction outwards as progressive
design and differentiation, could be written: 4 = 3 + 1, 3 = 2 +
1, 2 = 1 + 1…
Applied to the organism:
4. Systems →> organs +
"infrastructure": canals, streams, communication
3: Organs/cavities →>
tissues layers + channels, transport routes
2: Tissues →> cells + cell
projections and produced extracellular fibers.
1: Cell level with cell projections as structures or translated
into motions in dimension degree (d-degree) step 1-0/00,
designed in many ways at different levels.
The development takes the opposite way.
Links between steps according to the loop model:
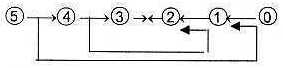 Fig
L-3 Fig
L-3
 Fig
L-4-29-3 Fig
L-4-29-3
2. Systems:
Nutrition and a sensory/motor system as
mutually opposed vector fields from V- and A-poles (see Embryology)
are both bi-directional in themselves, illustrated here as superimposed:
Inward/outward directions of vector fields
(d-degree 4):
a) Nervous system: sensory nerves inwards, motor nerves outwards.
(From ectoderm.)
b) Nutrition system: digestive organs inwards; blood system for
distribution outwards.
(From mostly endoderm.)
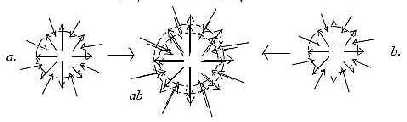 Fig
L-5-30-1 Fig
L-5-30-1
Real neurons are found first in Diploblastica, multicellular organisms
with sooner 2.5 layers (see Evolution)
but even unicellular organisms have sensitivity for light and excitation.
Nutrition system and nervous system are interwoven in such a fact
that the same peptides can be both digestive enzymes and neurotransmitters
in the nervous system.
However, the two systems operate mainly with chemical
versus the electrical one. In the dimension chain of physical quantities
mass and charge has been interpreted as a relation d-degree 3 to
d-degree 2. The neural system operates primarily with charge over
the surface of the cell membrane (d-degree 2), the nutrition system
with molecules representing what can be regarded as mass in this
context, in the interior of the cell (d-degree 3).
Hence, we can see the two systems meeting from
opposite poles in d-degree step 3 - 2
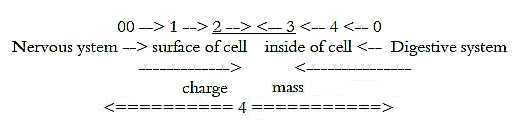
Fig L-6-30-3
Systems —> Organs:
Regarded as vector fields of d-degree 4 in our model, both primary
systems develop a growing complexity of double-directed infrastructures
(of d-degree 1 on a macro-scale). The vector fields get expressed
in d-degree steps ... ←
2←1←0/00.
Cf. 0 and 00 are "outer poles" or partial structures of
d-degree 4. Constructions get stepwise substantiated as from field
lines, from motional pathways to linear strings and canals etceteras...
(From steps 5 →
4 → 3 the steps
2 ← 1 ← 0/00 may be said to follow as debranched
d-degrees meeting in opposite direction according to the loop version
of the model, see figure L-3 above.)
Polarization of directions within the two systems:
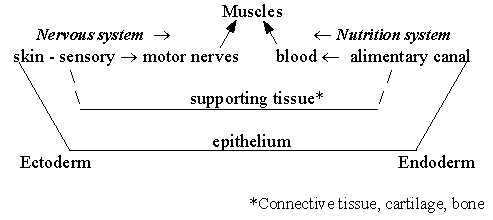
Fig L-7
- An essential part of body mass (d-degree 3) is made up of muscles
and supporting tissues as skeleton and so in, mostly from mesoderm,
the 3rd , middle tissue layer.
It could be mentioned here too that muscles
need innervation to develop.
3. Organs individualized:
Individual organs in relation to the "vector systems"
could be compared with word classes in relation to syntax in languages.
The stepwise synthesis from lower d-degrees 3←
2← 1←
0/00 can be referred to the integrating
force from 0-pole (cf. Exogastrulation),
while the individualization of centralized organs from the field
level can be suspected as an effect of the polarizing force of anticenter,
the 00-pole.
Organs, from the main "vector systems":
(The border between main "systems", "organs"
and "tissues" can be discussed.)

Fig L-8
Origin of the systems, approximate positions in blastula stage
of an embryo:
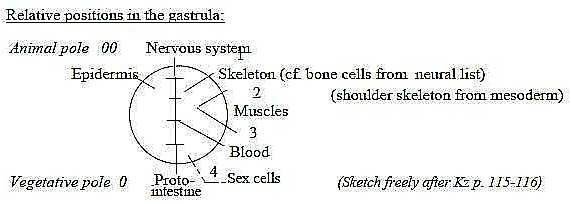
Fig L-9-31-3
With the assumed secondary developments of dimension
chains within each step of a basic chain, the individual organs
as more or less "tied off" and centered units (like organelles
in single cells) could perhaps be imagined as expressions for such
secondary evolutions designed in zstep d-degree 3 to 2. The principle illustrated with the unpaired organs
from the nutrition system:

Fig L-10-33-3
(Tonsils, thyroid and parathyroid glands and others
in the gullet wall not included.)
The animal pole and nervous system doesn't show a similar row of
secondary individualized units apart from the spinal cord and brain.
Cf. again Exogastrulation
in file Embryology,
Nutrition →> Blood
system:
The nutrition system develops a primarily "radial" distribution
system outwards, the blood system. A branched forefront of the intestine
can replace a blood circulation in primitive species.
In the embryo of less primitive species the blood
channels develop from archenteron as a dorsal and a ventral aorta,
along the F-B-axis. Blood is also in lower chordates produced by
the spleen, which in turn is derived from the intestine. And lungs
in the blood system develop from the gut.
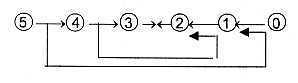
One example of the loop version of the model (see
figure 3 above) with debranched degrees from higher
d-degrees meeting 'the other way around' could be how blood later
gets produced in skeleton bones; skeleton as supportive tissue interpreted
as of lower d-degrees, see below about tissue levels.

Fig L-11-33-2
Nervous system →>
Muscles:
Development of muscles requires the nervous system (at least striated
muscles of vertebrates). Muscles atrophy if they are not innervated.
(The nervous system from the 00-pole in our application of the dimension
model, should as a first polarizing force appropriately act as a
differentiating factor.)
In the skin bag of "2.5"-layer species
(Diploblastica) sensory nerve cells and muscle cells are
differentiated adjacent to each other with parallelly running threads
in a layer of the skin bag, muscle filaments inside nerve fibers
(figure Ez p. 62).
It's perhaps noteworthy that these primary muscles
act circularly contracting and that the active force of muscles
is convergent, geometrical features derived from anticenter, the
00-pole in our model.
4. The organism as such and principle of individualization:
The demarcation of the multicellular organism by the skin, the
outer ectoderm, and its reproduction capacity through stored genetic
information in sex cells could be regarded as primary in relation
to development of the vector fields, as 00- and 0-poles respectively
in our model. They derive from opposite poles of the first embryo
and could perhaps be called the "individualization"
system.
The immune system with thymus, spleen and lymph
system is linked with the blood system and implies breaking-down
processes as phagocytosis in similarity with the inward directed,
digestive phase of the nutrition system.
Immune system could be seen as an expression of
the individualization principle on the deep, underlying level, as
a biological defense from inside the genetic heritage, developed
through the nutrition system. The skin is an equally essential counterpart
of an individual existence, an own(-ed) anticenter and a more mechanical
defense.

Fig L-12-31-2
5. Tissue level, 4-5 types:
Biologists talk about 4-5 tissue types, where the liquid one is
regarded as the 5th "tissue" (Mf, Kz).
Order of tissues according to their origin during embryo evolution:
Epithelium: appearing in ecto-, endo-, mesoderm
Nerve tissue:- from ectoderm
Muscle tissue: - from mesoderm
Supporting tissues: connective tissue, cartilage,
bone: - from mesoderm
Body fluids like blood, the 5th "tissue".
The tissue types can be described and ordered after degree of cell
contacts and of extracellular substance produced, i.e. different
degrees of internal versus external fiber production. It implies
a chain of decreasing "density" (in a general sense),
in this respect resembling the stepwise transitions in a dimension
chain from higher degree of internal structure to more externalized
relations and 1-dimensional forms as debranched d-degrees. (Cf.
density, according to previous definitions of physical
quantities the first "quantity".)
- Epithelium: Dense cell contact. Epithelium dresses
all outer and inner surfaces. It represents obviously the fundamental
form of 2-dimensional tissue surfaces.
- Nerve tissue: Contacts between cells through
their projections via synapses.
Single tubules of protein fibers within axons and dendrites.
- Muscle tissue: The cells separated by connective
tissue, cells = "muscles" almost completely filled with
protein fibers.
- Supporting tissues, reticular type: Cells more
or less scattered in the extracellular substance. When cell contacts,
these are just point-formed contacts between long cell projections.
External production of protein fibers as elastin and collagen.
With such fibers as 1-dimensional threads external
2-dimensional layers are built and 3-dimensional networks of external
relations.
- Liquids: No cell contacts. (Blood cells lose
also their nuclei.)
Both liquids and external fibers can be classified as secretion
products, analogous to "branched off" lower dimensional
degrees from higher dimensional steps in the dimension chain.
 Fig
L-13 Fig
L-13
The loop version can illustrate a connection between first tissue
type and the last one: epithelium forming glands with liquid secretion.

Fig 14-35-1
It's more doubtful if a similar relation may be found between nervous
and connective tissue in next step. However, we have osteoblasts
deriving from the area around the neural plate of blastula, among
other cells becoming e.g. glial cells. Osteoblasts form connective
tissue in bones, often in concentric circles around blood vessels.
(Perhaps also glial cells as astrocytes in the
brain can be regarded as "connective tissue", although
not described as such? )
6. Cell contacts:
Cell contacts are classified as follows (Kz) and show up
to be clearly 3- to 2- to 1-dimensional (possible to complete with
a 0-dimensional one in reticular tissue.):
 Fig
L-15-36-1 Fig
L-15-36-1
3-dimensional: Symplasma: nuclei of cells share plasma within
the same cell. There are two complementary types representing opposite
directions.
a) Plasmodium: Fission: a nucleus is divided into several
within a shared cell membrane; example (some?) liver cells. Note
outward direction: liver part of the blood system, subsystem to
the nutrition system.
b) Syncytium: Fusion of several cells to one within a common
membrane; example striated muscle cells. Note inward direction,
muscles as subsystem to the Nervous system.
2-dimensional: Wall-to-wall contacts, typical for
epithelium.
1-dimensional: Desmosomal junctions: long projections
of cells in wall-separated point contact; example reticular tissue.
Hence, contacts as common room, common wall, common creation of
"lines" through extensions but only a common "point".
We may note that in embryo development the first
divisions occur within the cell as mentioned in that file, which
corresponds to 3a here, plasmodium. Then, in the blastula, we get
the wall to wall-contacts. First during later gastrulation and from
mesoderm the 3b) type, syncytium appears. It seems to agree with
outward and inward (polarizing - synthesizing) directions in our
dimension model.
7. Fibers - levels of storage:
It is noteworthy that number of storing levels of extracellular
fibers at the final stages in the chain of tissue types are 5 -
4, from the individual molecules to the fiber - as if they corresponded
to steps in a chain (Kz p.154, 139, Fb p. 29):
- Collagen: 5 levels: collagen molecule - protofibril
- collagen fibril - bundle of collagen fibrils - collagen thread.
- Hair: 5 levels: keratin molecule - protofibril -
tonofibril - bark cells - hair.
- Cellulose fiber: 4 levels.
(Cf. chromosomes: 5 storage levels of the double helix of DNA.)

Fig L-16
To 04. Glands
|