|
1. Cell types in the nervous system (Ns):
Besides nerve cells there are several other additional cells around
them in nervous tissue, with a generic term called glial cells.
They correspond to anticenter in relation to central nerve cells
in a polarization of neural tissue, also representing higher dimension
degree (shortened d-degree) versus next lower one.
If on the cell level nerve cells illustrate d-degree
4 in our model, with their long axons and their dendrites (directions
outwards/inwards), glial cells will represent
3 (mass) and 2 (surfaces).
The glial cells derive from the neural crest, anticenter to the
invaginating neural tube.
They are further about 10 times as many, i.e.
make up most of mass in the brain, a multitude versus nerve cells.
This information concerns humans (Mf). (One hypothesis in
the dimension model has been that d-degree steps are connected with
10-power steps.)
Moreover, glial cells develop later in the fetus
and in the history of evolution. They are absent in certain simply
organized organisms, and myelin sheaths for instance, formed by
glial cells, are mostly missing in invertebrates (Fz).
Additionally, the relation between nerve cells
and glial cells is expressly said to be of the complementary type
in their internal processes: changes proceed in opposite directions.
Increase of a substance in the one type gives a decrease in the
other and the reverse (Kz p. 264, BA p. 115).
Apart from the immense amount of new knowledge, there are 5 types
of glial cells mentioned in older references here: 3 in central
Ns (CNS), 2 in peripheral Ns (PNS) and among these it's possible
to identify steps towards lower d-degrees.
Central types: astrocytes, oligodendrocytes and
microglial cells.
Central glial cells:
Astrocytes are star-shaped (fig, Nf p. 293) and have
functions and geometries illustrating both d-degree 3 and 2 in our
model with outer poles 4a - 4b of directions and radial and circular
poles of d-degree 2. Their extensions form" radii" (~
pole 3b) between nerve cells and blood vessels and are thought to
be responsible for transport of nourishment to the nerve cells.
Circularly (~ pole 3a) they tightly surround nerve cells. They are
a filling material with supporting functions and in addition make
up surfaces, lining all membranes and surfaces of mesodermal origin
in the brain. Astrocytes exist both in the gray and the white substance
of the brain.
Oligodendrocytes are mostly found in the white substance
of nerve fibers - as if representing a step-displacement from astrocytes.
They seem to enclose the nerve cells in a more ring-shaped way at
their extensions. They form myelin sheaths around axons in the central
Ns through fusion; cf. fusion as out of inward direction. It's a
polarity of the type nucleus - shell, a relation d-degree 3 to 2
in structure. Cf. about peripheral Schwann-cells below.
Microglial cells, the 3rd type in central Ns, have been considered
not to belong to the real neuroglial but to originate from mesoderm.
The cells are phagocytes and have an amoeboid mobility.
Thus, they seem to be a kind of wandering mesenchyme cells, the
final step in the chain of tissue kinds.
To summarize, these three kinds of glial in central Ns have features
that can be associated with last three steps in a dimension chain:
3 →> 2 →>
1→> 0/00.
Peripheral types of glial cells: Schwann cells and Satellite
cells:
Satellite cells are small cells whose short projections
surround the cell bodies of the big nerve cells: a typical anticenter
as periphery and also a multitude in relation to the unit of the
nerve cell (Photo Kz p. 162).
Finally, the Schwann cells form myelin sheaths around the
long extensions (projections) of nerve cells, the axons, in PNS.
In opposition to the cells that surround the very
nerve cell bodies in CNS and in opposition to the similar role of
oligodendrocytes that surround axons in CNS, these sheaths are formed
through a rolling up their membranes (d-degree 2) around
the axons as a kind of spiraling rotation. This opposition could
be taken as an example of how the 0-00-relation center - anticenter
changes character towards PNS and lower d-degrees: the rotational
motion as 2-dimensional an expression for debranched degrees in
lower steps.

Fig
Ns-21-85
They give also an illustration of how a magnetic field, surrounding
an electric wire, may be substantiated towards higher, superposed
levels. Or geometrically an analogy to this. The relation becomes
perpendicular as proposed in step 3 - 2, the radial versus circular
poles.
In their function it's possible to see a parallel
to the polarity inhibition - stimulation but rapidly repeated on
the same signal, a quantifying of a line. They maintain membrane
polarization, equivalent with inhibition, with nodes between them
for depolarizations, ~ stimulation. Also a form of pacing out a
distance as we have described the last d-degree step 1 →>
0/00 in the model.
And the physical quantity velocity
increases through this arrangement.
2. The nerve cell:
Nerve cells and sex cells are in certain respects opposites
as 00- and 0-poles:

Fig Ns-22-31-2
Sex cells have potential for maximal differentiation while nerve
cells are fully developed at start - and earlier thought not capable
to divide. Sex cells are haploid before fertilization, nerve cells
often tetraploid, so for instance in cerebellum.- a relation 1/4
in number of chromosomes.
The earliest nerve cell in history of evolution seems to have been
a combined sensory and motor cell, a sensory receptor cell with
motor axon. Indications of such cells have been found in the tentacles
of sea anemones for instance (Ez p. 385 f).
Then, the development has gone towards further
polarizations, a division of functions on sensory cells and motor
cells etc.
In the nerve cells from neural plate and neural tube the inward
conducting projections, the dendrites, are many, the outward conducting
projection one, the axon, as the 00-pole represent multitudes versus
unity of the 0-pole.
A nerve cell: dendrites and axon:
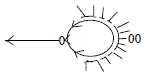 Fig Ns-23-87
Fig Ns-23-87
In the macrostructure of Ns it's the motor neurons that first gather
to centers in invertebrates - possible to see as an example of the
primary function of the 0-pole.
Arrangement of dendrites can have different structure
but is generally more or less circular around the cell body (inward
direction transformed to circular structure in 3rd d-degree according
to the dimension model). While the axon outwards branches radially
at target organs, an example of radial structure in d-degree 3 originating
from 0-pole in the model.
(Diameter of a nerve cell is about 5 - 100 μ,
the one of an axon about 1 - 20 μ
(LEL p. 27). Thus, the quotient should be circa
5/1.)
Other types of nerve cells are the bipolar and "pseudounipolar"
ones which develop from the neural crest, i.e. from anticenter in
relation to the neural tube.
Fig
Ns-24-88-1
These cell types are possible to interpret as secondary in relation
to the multi-dendritic ones. They belong to the inward-conducting
sensory, peripheral system, the bipolar one for instance found in
the retina.
The pseudounipolar type develops from the bipolar,
which can be regarded as expression for a center displacement, the
conducting fiber displaced out from body of the cell. Center displacements
towards lower degrees and higher levels are one principle in the
dimension model.
Thus, the series multi-dendritic →>
bipolar →> pseudounipolar
cells could be described in terms of angle steps of their extensions,
from 360° to 180° in/out to 90°
in relation to the cell body.
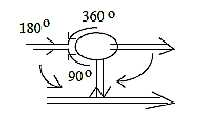
Fig
Ns-25-88-2
Nerve cells contain much of protein filaments and tubuli in the
cell body and out in plasma extremities, dendrites and axons as
well as in cilia.
Such organelles are common in other cells too as cell skeleton in
the cytoplasm. A coordination of motions is also found in unicellular
organism without a nervous system as protozoans (Ez p. 385)
- organelles with conductive ability.
Hence, nerve cells seem to be a specialization
in this respect of primary radial transport structures and vector
fields.
3. Nerve signals:
a) Two-way → one-way direction:
Some information indicates that nerve signals in an earlier stage
of evolution were two-way directed - first later become one-way
directed through chemical one-way direction over synapses. One has
for instance found a "mirror symmetry" over synapses in
jellyfishes with synaptic bladders on each side of the synapse (BA
p. 114).
In the early evolution of Ns dendrites and the
very membrane of the cell body seem to have had an ability to react
on electrical impulses, while they later only are chemically
excitable. According to another source (Ez p. 385) nerve
impulses in invertebrates sometimes seem to go in all directions
in diffuse nerve nets.
Such observations, if still valid, indicate an
evolutionary polarization from double-direction to differentiation
of functions and directions as in step 4 →>
3 →> 2 in our model. Cf.
a similar polarization regarding stimulation - inhibition above.
b) Two phases in signal transportation, chemical and electric:
The synapses could be described as a discontinuity, an "energy
gap" with a term from plasma physics. Such energy gaps should
correspond to a transition from one physical quantity to another
according to suggestions here, ultimately a change between d-degrees
in a fundamental underlying dimension chain.
The "carriers" of the nerve signal as
a force changes from electric to chemical to electric again.
From the viewpoint of biochemical
phases, defined by types of chemical elements and bonds,
the chemical phase with organic transmitters with bonds in 3 dimensions
may be defined as phase 3 in relation to the electrolytic phase
with metal ions, carriers of the electric signal.
Underlying these two phases we have the elementary
physical quantities Mass - Charge, assumed as a relation d-degree
3 - 2 in the dimension model.
D-degree 3 may be regarded too as a deeper, underlying
level, a binding force between charges on the superposed
level, appearing in the synapses. To compare with how hormones were
carrier of the information system before a nervous system developed.
(In the dimension model higher d-degree is defined as binding force
in relation to next lower one.)
In addition, transport of the transmitters occurs
in the center of axons, transport of charge along its surface, its
membrane, also showing on the polarity 3 - 2 with its roots in the
0-00-polarity.
c) Charge - electromagnetic waves (EM-waves):
Charge as a physical quality in d-degree 2 according to presumption
in the dimension model becomes connected with surfaces. An axon
of a nerve cell is excitable even without cytoplasm. The electric
potential should then be located to the border layer at membrane
(d-degree 2) of the axon (Zf p. 182).
The electric current follows from changes in the
voltage-potential over axon membrane, carried through by inflow
of Na+(sodium) and outflow of K+(potassium).
The outflow of K+starts
first about 0,5 ms after the inflow of Na+,
when this reaches its maximum. This "phase displacement"
resembles the one between electric and magnetic components in an
electromagnetic wave and could probably be interpreted as a related
formulation of the same structural principle - with K- and Na-ions
corresponding to E- and M-factors in an EM-wave.
d) "Motions to / from each
other" as poles 1a - 1b:
When Na+flows in through the membrane at
an impulse, it means a depolarization over the membrane to 0 as
in the dimension model "motions towards each other", pole
1a with origin in inward direction defines a 0'-pole. When K+
then flows out, it implies a re-polarization as "motions from
each other" (pole 1b) with origin in outward direction defines
an anticenter, a 00'-pole.
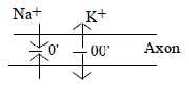
Fig
Ns-26-91-1
The signal propagates at straight angle to in- and outflows as
in EM-waves. Axons as lines are quantified.
Fig
Ns-27
e) How does the nervous signal propagate within the axons?
The answer seems not very clear. The transport of electric currents
is not depending on ion wandering in its cytoplasm. One theory is
that the "wave" propagates through displacements of charge
in a "bridge" of water molecules (Zf p. 202).
It's was said above that an axon is excitable
even without cytoplasm. In later sources (Wikipedia) the
"electrically conductive" cytoplasm is seen as explanation
for the internal spread of a wave from the local action potentials.
Myelin sheaths that inhibit in- and outflows,
increase the velocity of propagation (distance per second). They
increase length of the steps between nodes, the distance. Hence,
it cannot be transport of the ions in cytoplasm that drives the
propagation but some more immediate change of charge as through
electron displacements.
The action potential is transversal, the propagating "wave"
longitudinal. It's processes in different dimensions. We could think
about "waves" that in relation to "mass" may
be interpreted as a development - or aspect - in the less substantial
lower d-degrees 2 → 1
→ → 0/00.
In the series of chemical
forces as expressed in bonds we have identified ion bonds
in d-degree step 3 - 2, dipole bonds in step 2 - 1 and van der Waals
bonds in step 1 - 0/00. The nervous signal
appears as an expression for steps between these ion and dipole
forces (probably also van der waals moments involved).
The exchange of ions (Na+
/ K+)
with same charge seems driven by a concentration gradient, in terms
of density mass/volume, and becomes in
some sense a "binding force" as of higher d-degree in
relation to quantified "dipole" waves of charge as electric
currents. Mass versus charge in the model defined as a relation
d-degree 3 to 2 and higher d-degrees postulated as binding force
in next lower d-degree.
There is the step from ions as whole atoms to
electrons as carrier of the forces, and these simultaneously coupled
to a step in phases,
corresponding to d-degrees 3→>2 →> 1: crossing
of membrane as mass in relation to the more fluid medium inside
membrane.
Another feature is the step from two-way to one-way
direction, also an expression for a polarization step: An action
potential (the ion exchange) may actually give electric currents
in two directions, for instance when ignited at the middle of an
axon. The phase displacement between in- and outflows of the ions
becomes responsible for - or transformed to - the one-way direction
of the current.
Direction (d-degree 4 in the model) of the concentration gradient
and the electric current is closely connected, seems to guide all
steps.
Experiments show that if the polarity over the
cell membrane artificially is reversed to positive inside, negative
outside, the nerve impulse takes the opposite direction, backwards
in axons.
Expressed in terms of the dimension model it reveals
the interdependence between polarizations in different d-degrees
and poles that have the same character inherited from 0- or 00-pole.
When it concerns the chemical transport in the same direction within
the axons, one has also found a type of peristaltic waves, which
seem driven by the myelin sheaths. Cf. ring-shaped muscles around
intestines. Such a peristaltic wave with transversal, circular and
longitudinal contractions gets the same geometry on the chemical
level as the nervous signal on the electric level.
Fig
Ns-28-91-3
(Statically they get the form of standing EM-waves, cf. about worms
in Evolution.)
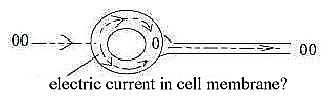
f) "Ignition" of the nerve signal - curious details:
According to general descriptions the frequency modulated impulse
gets ignited at outlet (hillock) of the axon from the cell
body.
One could possibly imagine the process as in this
figure:
Fig
Ns-29-92-1
However, it seems to be a curious fact that the action potential
gets ignited from a trigger point further out in the axon (Nf
p. 103).
It sounds as if there was some sort of an "imaginary"
quantum jump in the impulse through another dimension.
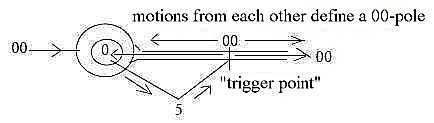
In the dimension model 2 poles of a certain d-degree
virtually exist as synchronous in next higher (underlying) d-degree,
before the step of polarization to the lower d-degree has released
1 d-degree as a new factor of distance or time (motions).
With the cell body as center, ~ a 0-pole, and
the axon directed towards anticenter, the 00-pole, some location
on this could be defined as a primary anticenter (decided by a distance
or quantified by some other factor).
It's then possible to think that the "quantum
jump" passes through underlying, next higher d-degree (as d-degree
5 in the figure or d-degree 1 in relation to d-degree 0/00
of motions): the higher d-degree that includes both poles?
Fig
Ns-30-92-2
Compare perhaps cilia
where the motion can start furthest out and not seems driven by
the cell body.
Another such curious observation: Out at sensory receptor cells
the inward conducting nerve cells seem sometimes to have synapse
vesicles towards the receptor cell, as if it sooner were the nerve
fiber that affected the receptor cell than the reverse.
This could perhaps be a related phenomenon, which
could be described as a "picking up of stimulation" -
from the opposite pole via another dimension, an underlying not
polarized phase or d-degree?
g) The action potential, levels and ion numbers:
A nerve impulse:
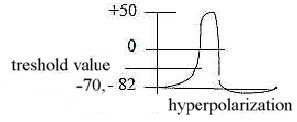
Fig
Ns-31-95-1
We can count on 5-6 levels of the potential over the membrane during
one action potential:
| |
Charge inside
|
| impulse maximum - point for change of directions
|
+
|
| 0 - zero charge - passage through the zero
line |
0
|
| threshold level - "angle change"
to "vertical" rise |
-
|
| | facilitated (graded) |
-
|
| normal rest potential - "horizontal"
level |
-
|
| hyperpolarization - "horizontal"
level |
-
|
A sketch on the steps as a whole dimension chain:
- depolarizations interpreted as steps inwards towards higher d-degrees,
- polarizations as steps outwards towards lower d-degrees in the
chain:
Steo 1 --> 0/00 debranched from step 5 --> 4:
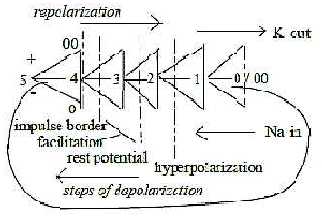
Cell memrane in step 3 - 2.
Inhibition -- Stimulation 2> 1 < 2
Amplitude modulation in d-degree 3.
Impulse trigger in step 3 - 4
- Impulse between outer poles 0 -- 00.
Fig Ns-32-93-1
Ion numbers:
Na 11 Z, 3rd shell, as ionized 10 e-. 2nd shell
K 19 Z, 4th shell, as ionized 18 e-, 3rd
shell
Numbers in the 2x2-chain behind the periodic system:

Fig
Ns-33
Both atoms in the s-orbital of the shells, representing step
1-0/00.
Sum Z of Na + K = 30 = 2 times the sum of an elementary chain 5
→>
0:
15 -/+ 4 = Na Z - K Z.
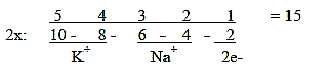
Fig
Ns-34-94-2
Concentrations of K+ and Na+
ions inside - outside the membrane in a certain part of the axon
(1 μm x 1 μm x 0,1 μm
according to reference Mf p. 35:
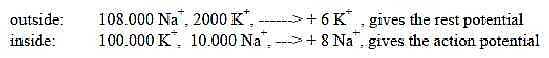
Fig
Ns-35- 94-3a
Since the sums happen to give the sum 110 x 103 of the
2x2-chain above, the figures could be illustrated in
this chain:

Fig
Ns-36-94-3b
Cf. steps 1 - 2 - 3 and number relations between Na- and K-ions
that get pumped in and out by the Na-K-pump to restore the rest
potential concentrations. In different studies and different cells
it has been found that the relation Na /
K can be 2:1 or 3:1 or 3:2 (Nf. p. 43)
To Nervous
System: Brain parts
|