|
1. The basic units of muscles:
Muscles are in the service of locomotion, both internal and external.
How to interpret this fundamental and intricate invention?
Threads of F-actin are found in most cells
and F-actin is said to be among the most common proteins in animal
world.
Cells
include a rich net of tubuli constructed of protein spirals
from the membrane of nucleus outwards as a more or less complicated
vector field transformed to radial and circular structures. (The
nucleus then as a complex 0-pole.) They seem to function as ways
for transportation of substances. So also for instance in axons
of nerve cells. Such tubuli become the essential components in cilia
and flagella, the first organelles for external locomotion of unicellular
organisms. The contractile proteins in these have also been described
as a kind of actomyosin complex (Zf p. 212). (Many of the
involved proteins have been identified during later decades.)
The basic units of muscle cells within Z-bands of
a sarcomere are the threads of F-actin and myosin. They are
arranged in a way that in many respects illustrates the dimension
degree (d-degree) step 1 →>
0/00 in the dimension model, polarizations
of d-degree 1 to "motions from each other" (pole 1b) and
"motions to each other" (pole 1a). D-degree 0/00
in the model is defined as the d-degree of motions.
 Fig
M-1-54-2 Fig
M-1-54-2
- The linear threads (d-degree 1 on one level) of F-actin
and Myosin within the Z-bands appear as a polarization
in positions and motional directions:
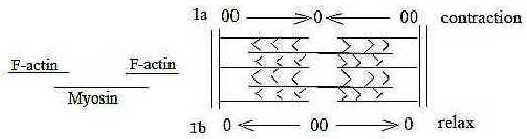
Fig M-2-5-1
- Myosin threads illustrate the "motions from
each other"; they are bipolar threads with "heads" outwards
and connected tails as center; the "heads" move outwards
in opposite directions when active.
- F-actin threads illustrate the "motions to
each other"; they are divided, the parts attached to opposite
borders of the sarcomere, an anticenter arrangement, and move or rather
get moved in direction inwards during active phase, returning to
original position during relax.
- It should be observed that it is Myosin, representing the center,
that is the active force, divergence (Vdiv), as 0-pole is defined
as the binding, integrating force in our model.
- The polarity between F-actin and myosin appears also in the
different construction of the threads:
Myosin (the thicker threads) is composed of double
spirals of simple protein chains, while F-actin is chains of globular
proteins (G-actin) as strings of beads. It could be regarded as
a relation between d-degrees 4 and 3 on the level of individual
proteins: vector-shape of d-degree 4 (myosin) versus the 3-dimensional
globular units of G-actin, representing quantification.
( Higher d-degree is defined as binding force
in next lower one in the model. So is myosin in relation to F-actin
in active moments.)
We have something of a similar relation between "linear"
arranged nucleotides in mRNA versus globular ribosomes, the
rRNA at protein
synthesis: a similarity perhaps worth reflecting more
upon.
- Further, F-actin contains about 300-400 G-actin and
the bundles of myosin about 300 parallel threads (Aph): hence
approximately the same numbers but F-actin lengthwise, myosin "vertically",
another complementary polarity.
If we regard the relation myosin - F-actin as a d-degree step, the
one d-degree debranched in such a step could be identified with
the transverse connections as "catches",
polarized into motions.
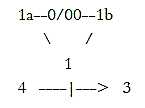 Fig
M-3-56-1 Fig
M-3-56-1
In the active moment myosin and F-actin get united through catches
on myosin which climb stepwise on actin in opposite directions outwards,
which results in the passive contraction between the two F-actin
halves. That's how the real force of activity originates from the
outward direction and is depending on center as binding force.
(Elastic thin protein threads of titin
keeps myosin bound to Z-bands.)
- Actually, the arrangement could in essential respects illustrate
human walking on a much superposed level, with F-actin
as the ground, bonds to active sites on F-actin as footholds. .
Fig M-4-56-2
There is the divergent moment, (like one foot forward) where the
head of myosin with attaching catches is pointing outwards, then
the drawing moment of convergence when the head of myosin gets more
and more inward directed until it loses the grip (like lifting the
back foot) - and a new moment of divergence starts. (A bit unfortunate
to call the catching place "heads" of myosin!)
Human beings have too legs and feet. The single
myosin is composed by two protein threads and has two heads as feet.
(Said with reservation for any parallel in interaction.)
In the dimension model we have that pole 1b (here myosin) as "motions
from each other" defines distance as a new anticenter pole
00' while pole 1a - " motions to each other"- defines
a new 0'-pole. Even these definitions seem applicable in the process.
Walking includes stepwise, a bit wavy displacements of the center
of gravity of the body. The bond of catches to active
sites of F-actin could imply such a displacement of center. The
process expressed in d-degrees should probably be described as going
via d-degree 1 as binding force of motions: 1b →>
1 →> 1a, 1a →>
1→> 1b.
(With the view on F-actin as representing the
00-pole, in the model primary polarizing force, we could suspect
some force from F-actin threads - or from borders of sarcomeres
as anticenter - on the myosin threads too, indirectly affecting
its divergence? The complexity increases when the analysis deepens
towards more microscopic, underlying levels and several other
proteins are involved. )
In fact, we could regard the walking or
the activity of muscle filaments as a kind of substantiated L-waves,
the type that hypothetical gravitational waves are assumed to belong
to. Two parallel, phase-displaced ones - or a polarized one!
00→>
0←00
→>0← 00 →>0 ← 00
→> 0 ← 00
→>0← 00 →>0 ← 00
→>0...
An additional question is if the relations myosin - actin eventually
could tell us something interesting about the relation between
the outward acceleration force
(FA) and gravitation (FG)
in macrocosm? In which sense could FA
contribute to the contraction (FG) of
masses?
- The structure affecting the binding moment is complex (Aph).
F-actin is in reality surrounded by winding threads of tropomyosin.
Each of these covers 7 active sites on F-actin. (Cf. earlier arithmetical
speculations about number 7 in file
Biochemistry, 1/7.).
At the middle of these are small molecules of
tropin attached, which have 3 binding sites: one to G-actin,
one to tropomyosin and one for Ca2+ that
induces the uncovering of active sites on G-actin, which get the
catches of myosin to bind to them.
In this sense the active moment starts from F-actin
and from Ca2+, a metal ion representing
the 00-pole among elements,
acting as such polarizing when breaking up the cover of sites on
actin.
If we as above regard the structures of myosin
and F-actin as a relation d-degree 4 to 3, could we eventually associate
the numbers of binding sites, divided in 7 on G-actin covered by
tropomyosin, and 3 on tropin, with these figures: 7 = 4 + 3, 3 the
d-degree of globular F-actin?
A note
An association, however wild it may be, about molecular weights:
The relation between molecular weights of myosin (~ 500 x 103)
and G-actin (46 x 103) is 500 to 46: F-actin as n x 46.
An association to numbers in the genetic code:
In the suggested number series behind the genetic
code, the numbers 500 and 460 appear, see the figure below.
And total sum of the 20 + 4 double-coded amino acids for 24 differentiating
codons = 6 x 546.
Ca2+, 40 A, is the ion that when attaching
head of myosin triggers a contraction.

Fig M-5-56-3
[Since mass number 101 of side-chain of Arginine represent last
step 1 - 0 (+1) in this exponent
series, one could wonder if this amino acid eventually plays
an important role in the structure for locomotion. It's said to
be rich in fish sperm-tails and is essential in the creation of
creatinephosphate and in the corresponding energy-storing substance
arginine phosphate among shellfishes.]
2. Levels of storage:
The number of levels from the single protein threads of F-actin
and myosin to a whole muscle is about 5 to 6, as number of steps
or "borders" in a dimension chain.

Fig M-6-54-1
Compare number 5 in levels of storage in other structures too as
those in a hair or in collagen.
[It's natural to ask how then the number of something
would be related to a chain of different dimensions. Through
a dimension chain as process outwards a 5-dimensional unit gets
translated to 5 x 1 linear dimensions. The addition of units
in each step inwards could be suspected as guided by convergence
from the 00-pole at end of the chain. (Just one aspect among
others. Cf. 5-merous flowers and fingers of a hand.)]
3. Muscles in chains of organs and tissues:
Muscles originate from mesoderm and can be regarded as the essential
intermediate organs between the two vector fields (file Levels),
the nervous and nutrition systems (as between poles 4a - 4b in the
dimension model). Striated muscles need innervation to develop and
one primary function of locomotion is seeking for food, demands
from the nutrition system (4b),
Muscles become massive organs and fill a big part
of the body between central intestines and the body surface of an
early chordate as the lancelet. In grown up humans the muscles make
up about half the weight.
In the chain of tissue
types, with regard to degrees of internal to external fiber
production, muscle tissue appear in the middle, from mesoderm but
still without the external fiber production characterizing bone,
cartilage and connective tissue.
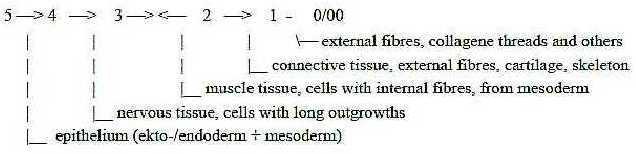
Fig M-7-47-2
A muscle as organ is encapsulated by connective tissue
and a web of reticular threads (steps 2 - 1 in the tissue chain),
while nerves and blood vessels (regarded as vectors from higher
systemic degree (poles 4a and 4b, outer poles of d-degree 3 in the
model) enter at straight angles through the covering, both through
outer perimysium and inner endomysium. Often together
(thus not as in the figure below).
Fig M-8-47-3
4. Nerve cells - muscle cells, evolutionary aspects:
A motor nerve cell has long axons with tubuli of proteins radially
outwards from the cell.
A muscle cell in an early version as in cnidarians
is the epitheliomuscular cell (Ez p. 59). It has the
protein fibers in straight angle to a short projection of the cell
and is part of a ring-shaped contraction system (Ez p. 59).
Hence, it resembles in structure a bipolar cell in the sensory nervous
system.
As motor and sensory pathways represent outward
and inward directions respectively and simultaneously have the relation
between higher and lower d-degree, so could this early muscular
cell reveal the relation between the nervous system and muscles
as organs. Cf. the angular step 180° to 90°, corresponding
to a step 4th to 3rd d-degree according to assumptions in this model..
 Fig
M-9-48-1 Fig
M-9-48-1
Muscles develop in similarity with other organs from lower towards
higher d-degrees: from muscle threads (as d-degree 1) to layers
(2) to muscle masses (3).
In the history of evolution first muscles seem to appear in
"2.5"-layer organisms, Diploblastica, with 2 layers
and an intermediate substance but no real mesoderm. The mentioned
epitheliomuscle cells form a ring around the border of the bell-shaped
jellyfishes for instance. (However, already 1-layer organisms have
certain cells that resemble visceral muscle cells (Ez p. 54).
Later during evolution a skin-muscle sac develops
as for instance in earthworms: a tube-shaped layer of circular and
longitudinal muscles, which becomes one expression for poles of
d-degree 3 in our model.
Then, in 3-layer animals comes the development
of striated muscles out of the inner, thickened plates of somites
and several muscular layers combine to "muscle mass".
It would be a possible alternative to regard the design of muscle
tissue also as substantiation of external motional
moments in different d-degrees:, in this case from higher
to lower ones. As suggested in the model:
- In D4: Vibration, halved, just one of the moments, contraction,
designed in operation of the individual actomyosin threads.
- In D3: Rotation, motion in the plane: designed as a
ring of muscle cells.
- In D2: Motions in 3 directions, in one form rotation
plus "translation": designed e.g. in the tube-shaped
layer of the earthworm or e.g. in the peristaltic motions of intestines.
Note that these latter muscles in reality go in spirals (Kz
p. 187), another form of 3-dimensional motion: rotation superposed
a linear pathway. Or, as "translation in 3 directions"
(with the terminology from motions of 2-atomic gases) the 3 directions
of muscle fibers in the tongue.
- In D1: A 4-dimensional motion suggested identified with
pumping inwards / outwards: designed
in the heart sac and its spherical muscle net (also with a kind
of spiraling layers) and with internal radial muscles to valves
in the ventricles.
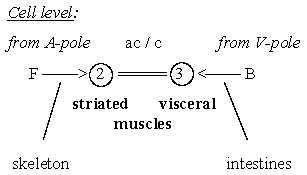
5. Muscle types:
a) Striated - Visceral muscles:
This polarity reflects both the original center - anticenter
one of vegetative - animal poles and the derived front - back polarity:
- Visceral muscles serve the central nutrition system, alimentary
canal and its glands, blood vessels - and womb (uterus), also an
organ for nutrition.
- Striated muscles connect (via tendons) parts of skeleton,
identified here as developed in d-degree 2 - 1 in the dimension
chain of organs.
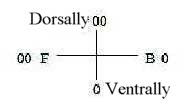 Fig
M-10-50-1 Fig
M-10-50-1
- The inner (splanchnik), obviously more ventral back part
of lateral plates of mesoderm gives visceral muscles.
- The inner part of the front, dorsal plates from somites gives
striated muscles.

Fig M-11-47-1
"Striation" seems to be an effect of the front and anticenter
00-pole. Visceral musculature around front part of the alimentary
canal during evolution develops to the striated type in jaws and
gill arches. And as mentioned above, (at least?) striated muscles
need innervation from nervous system as 00-pole to develop.
 Fig
M-12 Fig
M-12
The two kinds of muscles show on the cell level a polarity of the
type 0 - 00 in more than one respect:
One is the opposition unity - multitude, another center - anticenter:
- Visceral muscles have only one nucleus.
- Striated muscles have several nuclei. These are also result
of fusion, in this sense of inward direction, as from the 00-pole,
the front pole of the F-B-axis.
(There exist also a type of intermediate multiunit
smooth muscles (Aph) and in striated muscles single-nuclear
cells surrounding the other that have the ability to fuse, i.e.
converge from anticenter, and at damages begin to produce fibers.)
- Inner and back visceral muscles around intestines have their nuclei
in the center.
- Outer and front striated muscles have the nuclei at the periphery,
the anticenter.
- Visceral cells have filaments and fibrils unordered in different
directions.
- Striated muscles have the filaments paralleled and arranged in
the "sarcomeres", described in No. 1, and in myofibrils.
They also differ a bit biochemically in effectuation of the motions
(Aph).
How should this difference be interpreted with
aspects from the dimension model? We could see the "unordered"
multidirections of fibers in the visceral type as of higher d-degree
in relation to the paralleled, linear order and one-way direction
of the striated ones. More "order" towards lower d-degrees?
Cf. that striated muscles, gathered to the 4th level of storage
(fascicles) can have different directions: convergent, pennatem,
circular besides the parallel one (Aph).
- Visceral muscles have a certain own capacity of electric conductivity
- and are regulated by the deeper, autonomous nervous system, while
striated muscles are governed by more superposed centers in the
brain. Also, naturally, connected with the fundamental 0-00-polarity.
The innervation of visceral muscles differs too,
possible to regard as a geometrical expression for a radial 3-dimensional
structure versus a plane 2-dimensional formation:
- Visceral muscles have more unordered free nerve ends in the neighborhood.
- Striated muscles have end plates of the nerves in close contact
with the muscle cells.
b) Electric tissue:
Relative to other tissues and organs muscles have here been regarded
as representing a development in d-degree step 3 - 2. Thus, it's
interesting that muscles can be specialized to electric tissue.
With the general assumption in the model that Charge
in relation to Mass as physical qualities may be analyzed as a relation
of d-degree 2 to 3, this specialization implies a step or half one
to d-degree 2: the muscle fibers become flat layers, one-sidedly
innervated, stored in Volta piles with the ability of electric bursts.
(Simultaneously with a reduction of fibers.)
c) Heart muscles:
The heart muscles represent a third type besides visceral and striated
ones, combining properties of both, a kind of pole meeting between
the other types (cf. figure 12 above). They serve the nutrition
system but is positioned in front part of the body (~ that of the
00-pole), on an early stage in suprasternal fossa, the juggular
notch (!).
- Cells have only one nucleus, with central position as in visceral
musculature.
- Filaments are however striated, paralleled, but cells also branch
to a network; cf. radial divergence as one pole of d-degree 3.
Further, the musculature is autonomous in nervous activation -
as to a certain degree the visceral muscles. It has its own center
for nervous impulses. With the loop version of our model where opposite
directions meet in the central step 3-2, a new kind of center may
be assumed to develop. (See the Blood
system.)
Heart muscles in 3 layers form the "spherical"
sac, and inner radial muscles guide the valves; hence illustrating
the geometrical polarity circular - radial in d-degree step 3 -
2.
Radial components depart from a center. In similarity
with the radial muscles to the heart valves, departing from bottom
of the ventricles, which represent outward direction of the blood,
the signals from atrium go first to the bottom of the ventricles
before they spread upwards = outwards in the walls of ventricles.
6. Arrangement of striated musculature:
a) Limbs:
Striated muscles in the limbs get the function of binding forces
between skeleton parts as higher d-degrees in relation to lower
d-degrees in our model. They are principally "phase displaced"
in relation to sections of the skeleton bones in arms and legs -
in a certain similarity with the arrangement of myosin and actin
in the individual muscle cell fibers, - and with steps in walking:
 Fig
M-13-52-1 Fig
M-13-52-1
Since the activity of a muscle is only contractive, the polar function
between stretching and flexing moments (← → and →←) in motions is attained through complementary, antagonistic
arrangement of the muscles.
The upper arm of a human being taken as one example:
It has muscles with 5 origins as attachments, divided 3 and 2, triceps
and biceps.
- Biceps with 2 "heads" or origins is a muscle
on ventral side of the upper arm.
- Triceps with 3 origins is a muscle on dorsal
side.
This arrangement seems at first to contradict what we could believe
from aspects on the dimension chain where dorsal side originate
from 00-pole and lower d-degrees, ventral side from 0-pole and higher
d-degrees. It seems also to contradict biological needs; 3 origins
of a muscle should give more strength, better needed in flexing
activities of the arm than in stretching it? (Demonstration of strength
shows the biceps, not the triceps!)
The explanation seems to be just geometrical,
the arrangement founded in geometry: The 3-headed muscle stretches
the arm, expressing the radial factor of d-degree 3 in our
model, while the 2-headed biceps bends the arm, expressing
the circular factor of d-degree 3, derived from inward direction
and anticenter in the model. (Such facts could support the views
here that fundamental geometries rule.)
(Yet, the 2 origins of biceps attach to front parts of the shoulder
blade (scapula) on dorsal side of the trunk. (Insertion on
forearm, inside of elbow.) Triceps: 1 origin on lower part of dorsal
scapula, 2 origins on upper arm. Insertion on forearm, outside of
elbow.)
A muscle is mostly firmly anchored in one point, movable in the
other, which implies that it has a kind of direction. Cf. poles
4a-4b as outer poles of d-degree 3 in our model.
The complementary relation in the function of
these muscles appears also in lengths of their tendons: biceps has
long tendons proximally, short distally, while triceps has short
tendons proximally, longer distally (Mf p. 81).
Number 5 in origins of these muscles return - naturally? - in the
5 spinal nerves that innervate the arm. Possibly too connected with
the number 5 in hands and feet.
(The bending of the arm could also be regarded from the aspect
of angle steps, 180°→>
90°→> 45°...,
hypothetically assumed connected with d-degree steps 4 →>
3 →> 2...)
Muscles in the legs show the same agreement between number of origins
and geometrical function as the arms: the "4-headed" muscle
stretches the leg at the knee; a 1-2-headed muscle bends it. In
the legs however, the number of origins don't contradict the dorsal-ventral
polarity: the 4-headed muscle is ventral, the 2-headed muscle dorsal.
This fact could in itself follow from origin of
ventral and dorsal sides from primary vegetative 0-pole versus primary
animal 00-pole: arms at front part from original 00-pole, legs at
back part. It could indicate the reversal of reversal in directions:
0→> 4→>
3 →>← 2← 1←
00.
About the number of heads of the muscles, we could here remember
the loop version of the model which implies three steps of polarizations
of number 5:
5 →
0 + 00, 5 → 4 +
1, ( ~ legs), 5 → 3
+ 2 (~ arms).
Cf. legs — corresponding
to the d-degree step 4 →
3 from A-V-axis to front - back axis F - B. (See Embryology.)
b) Musculature of the trunk:
Along the median line of the trunk there are no muscles, probably
depending on the fact that muscles originate from mesoderm and somites
laterally, on either side of the central coordinate axis of the
gastrula.
Both longitudinal (rectus), transverse and oblique
muscles develop in the trunk, as expressions for angle steps 180°
→ 90° →
45° (associated with coordinate axes 4 - 3 - 2 in the assumption
about angle steps).
(The six outer muscles of the eyeball operate
in a similar way: horizontally, vertically and in oblique directions.)
The transverse muscles lie under the layer of oblique and longitudinal
ones as the ring-formed muscles around intestines inside the longitudinal.
It's naturally explained by the function: the latter should of course
not be included in contraction of the ring-formed, transverse ones.
It could however simultaneously reveal that the ventral - dorsal
coordinate axis in the plane of these muscles precedes the front
- back axis in the embryological
development.
Then, the two longitudinal, ventral muscles go closest to central
F-B line of the trunk, on each side, as in agreement with the dimensional
interpretation. They reveal a reminiscence of segmented animals
and segmented somites of the embryo in being divided by transverse
bands of tendons into 4 -5 or 6 sections ("the six-pack").
Notice again the number about 5 as steps in the dimension chain,
Muscles of ribs:
Inner and outer layers of oblique muscles develop more laterally.
Cf. the lateral coordinate axis L-R as the 3rd one in embryological
development, assumed as representing d-degree 2. After a d-degree
step 4 → 3 (180° →>
90°) we get a polarization in two directions to +/-
45° as illustrated in the figure below. This could explain why
there are two layers of oblique muscles and how one of these layers
on illustrations seems to lie under the longitudinal one, one outside
it.
 Fig
M-14-53-1 Fig
M-14-53-1
One definition of geometrical poles out of d-degree 2 in the dimension
model is the polarity inside - outside, with origin in the poles
outwards - inwards of d-degree 4.
The oblique muscles of the ribs that serve the breathing illustrate
these polarities:
- Inner rib muscles when contracted give exhalation,
~ outward direction of air.
- Outer rib muscles when contracted give inhalation,
~ inward direction of air.
Further, direction of the muscles at exhalation is outwards from
central line viewed in direction back →>
front, while direction of muscles for inhalation is inwards the
middle:
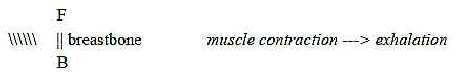
Fig M-15-53-2
Fig M-16-53-3
To 07. Skeleton
|