|
1. Ventricles and the CSF-liquid:
In the evolutionary development of the brain the
ventricles and their shapes could have been an equally decisive
factor as the development of neural mass, this with dimensional
views applied to the design. Compare mass - space as poles of d-degree
3 with neural mass versus ventricles.
Evolutionary development of lateral ventricles in forebrain, cross-section,
left and right cerebral hemisphere:
A simplified sketch after Kz p. 250-251:
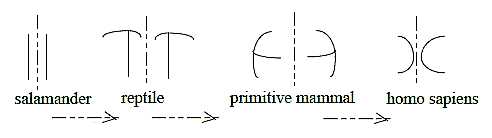
Fig
Ns-37-97-1
- Salamander: ventricles along longitudinal axes of the body - as
d-degree 4.
- Reptile: a polarization towards perpendicular relation through
a step d- degree 4 →> 3
(radial - circular as geometric poles of d-degree 3).
- Primitive mammal (Solenodons, a kind of mouse): whole ventricle
turned 90o - as through next step 3 →>
2.
- Humans: an inversion convex - concave of the half circles - as
in a d-degree step 2 - 1.
(It could be said that shape of the hemispheres
simultaneously develop towards increasing number of "sides",
from "two-sided" in salamanders, "3-sided" in
reptiles, "4-sided" in Solenodons to approximately semispherical
in humans.)
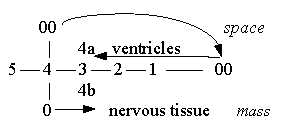
The ventricles 4, 3 and 2 in humans
has shapes that roughly illustrates the geometries of corresponding
d-degrees in the dimension model - with increasing size:

Fig
Ns-38- 97-2
- 4th ventricle: along the axes F-B, front-back, a widened canal
with a little polarization dorsal - ventral direction as between
poles 00 - 0.
- 3rd ventricle: a widened space volume.
- 2nd ventricles, division in two, bows surrounding 3rd ventricle
as "shells" around a central room.
Related parts of neural brain:
- Hemispheres of cerebrum around the 2nd ventricles.
- Diencephalon around the 3rd ventricle.
- Mesencephalon around the canal between 4th and 3rd ventricles.
- Medulla oblongata with pons on ventral side and cerebellum
on dorsal side of the 4th ventricle.
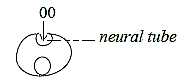
The central canal of the spinal cord as a cavity
is at start of the embryological development a built-in surrounding,
an insubstantial space, which widens and develops into design of
the ventricles, of the embryo, i.e. anticenter at the animal pole
that gets enclosed when the neural plate invaginates to the neural
tube, in positions a pole exchange ac - c, mass - space:
 Fig
Ns-39-098-1 Fig
Ns-39-098-1
The spinal canal with CSF becomes the opposite pole to the neural
mass around it. As a primary anticenter it could perhaps be suspected
that the CSF chemically induces such polarizations as for instance
the transverse bands on the spinal cord and divisions in brain bladders
(?). Cf. 00-pole as a first polarizing force in the model here.
While the canal with CSF develops stepwise to the central ventricles,
it takes a side-way too at 4th ventricle to circulate anticentrically
around the whole brain, a polarization c - ac in relation to inner
ventricles.
It's notable that this branching occurs in the
4th ventricle and in its ceiling and dorsally (the anticenter
side deriving from first animal pole) and through 3 small holes
(foramina). It illustrates how a step from d-degree 4 to
a geometry of d-degree 3 can be expressed biologically.
We have in the dimension model that the 00-pole
may be regarded as debranched, meeting "the other way around"
in a haploid chain, inwards to circular pole 3a:
 Fig
Ns-40-98-2 Fig
Ns-40-98-2
Another expression for the primary character of inward direction
of CSF is that it's produced in inward - backward direction from
the ceiling of 3rd ventricle, the floor of 2nd ventricles (hence
in step 3 - 2).
It is non-neural tissue epithelium, dorsally in
the neural tube in forebrain and diencephalon that has been displaced
inwards towards the inner ventricles and has become gland epithelium
for the production of CSF (Kz p. 242). Hence, in several
respects expression for anticenter and inward direction.
In the relation neural tissue - CSF we have naturally also a relation
between phases,
between organic matter and liquids of the kind that can be described
as 3- to 2-dimensional with regard to chemical bonds. (Content of
proteins etc. in the liquid here neglected.)
CSF contains more of Na+
ions than other extracellular liquid, which could be a sign of its
origin from outer surrounding of the embryo?
The relation CSF-canal —
surrounding neural mass can be compared with the elevator versus
stairs in a building (possibly both chemically and psychologically).
Also a relation continuum - quantum jumps.
"Reissners thread" is a mysterious line
of glycoproteins with unknown function that goes from secretory
cells in diencephalon backwards through the whole CSF-canal. It's
believed to serve transport of molecules. It could be regarded as
one expression for the main coordinate axis F - B. Perhaps it corresponds
to "the other way around" in the figure above! It's only
absent in primates, which eventually could have connection with
the loss of a (real) tail?
(It seems as if a part of this fiber (RF) could
have an impact on outgrowth of axons in e.g. chicken embryos (http://www.springerlink.com/content/n80v070740616q62/).
According to information some decades ago it seemed to be internal
secretion of substances from the circulating CSF-liquid into the
brain that induces sleep,
among other molecules GABA (γ-Aminobutyric
acid). Together with the neural center for sleep in medulla oblongata,
it illustrates the double-direction 00 <=====> 0 inwards-outwards
during sleep: the phase for animals as "whole worlds"
or entities in themselves, both centers and anticenters.
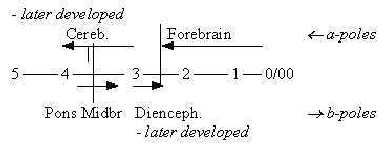
2. Brain parts:
§It can be noted that the number of bladders on front part
of the neural tube that develops to a brain is 5 already on an early
evolutionary stage of craniates, 5 with the widening of medulla
oblongata: in number corresponding to steps in the dimension model.
After a rearrangement to 3 and new differentiations
the bladders become these 5 well known parts of a human brain:
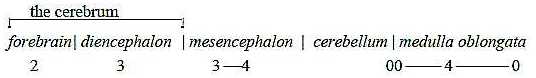
Fig
Ns-41-99-1
(Figures are preliminary identification of d-degrees,
more closely commented below.)
The long evolution of the nervous system is similar to other organs
as the blood system: a development 0 →
1 → 2 →
3 in dimensions from single neurons to a "linear" tube
to wavy forms with curves (concave - convex) to a more and more
centralized mass.
From spinal cord to the brain there is a rearrangement
of neurons from the "linear" order in columns to an arrangement
in separate, more centralized nuclei, also a dimensional development
towards d-degree 3 on the neuron level.
The prolongation of the neural tube to the brainstem can be regarded
as a center in the centralized mass of the brain - with root in
the body.
A general principle is that integrating centers
lie deeper down with increasing differentiation outwards the surfaces.
a) Medulla oblongata and Pons:
Some of the functional centers mentioned below are actually located
to Pons on the ventral side of medulla (Wikipedia and earlier
sources).
Medulla oblongata around the 4th ventricle contains the reticular
activating system RAS (ascending part ARAS) - with a general, unspecific
and divergent spread of pathways upwards to the whole cortex for
an arousal level and downwards for muscle tonus for instance. The
neurons have extra richly branched axons.
These systems become in the interpretation here
an obvious expression for d-degree 4, the level of vector fields.
Centers for consciousness as such are located here
but also for sleep, playing a role in generating dreams.
Cf. sleep as pointed to above is a phase of two-way direction, d-degree
4 as only "virtually" polarized.
Another example of the two-way direction is the
center for respiration that regulates the breathing:
in agreement with most other polarities inhalation ~ inwards:
dorsal part of the center, exhalation ~ outwards the ventral part.
Further, it's notable that the reticular formation
contains centers for the basic directions in postures
of the body and its balance around the center of gravity.
It gives another example of the 4-dimensional character of this
system.
The extrapyramidal tracts are chiefly found in the
reticular formation of the Pons and Medulla. It's essential to underline
that these tracts also have the ability to execute motions governed
by the will (LEL p. 160). A will that comes from direction
in this deep structure.
Furthermore, medulla includes centers for control of blood circulation
and elementary digestive functions, thus also for the vegetative
system.
Several other sensory and motor centers are mentioned
in Pons as for hearing, taste, eye movements and facial expressions.
In the "reticular formation" white and gray substance
is not yet separated as in the forebrain but a network of closely
integrated neurons and nuclei. Cf. polarizations as d-degree steps
outwards the forebrain on this macro-scale on the tissue level.
It's also interesting - and not astonishing - that some relationship
between RAS circuits and pathways for physiological pain has been
found. Cf. about pain
as one of the oldest and most general senses.
A last notice: The cranial nerve that governs the motor activity
of the tongue emanates from a location furthest back in medulla
oblongata. Cf. the connection between d-degree 4 in our model and
d-degree 0/00 of motions with speech as
the last in a dimension chain of psychological
faculties.
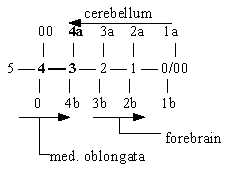
b) Cerebellum:
It's original and basic function seems to be a sensory integration
(pole 4a, inward direction in our model) when it concerns body positions
(in 3-dimensional space) and motions, thus in agreement with its
dorsal (~ anticenter) position. Cerebellum has also by several scientists
been regarded as at bottom a sensory organ (Wikipedia): it
receives the lot of inputs from both cerebrum and sensors from muscles
in the body and has regulating, inhibiting functions.
The smallest region, the flocculonodular lobe, is mentioned
as the oldest part in evolutionary terms, participating in balance
and spatial orientation. Its primary connections are with the vestibular
nuclei, the organ
for equilibrium, although it receives visual and other sensory
input too. Damage to it causes disturbances of balance and gait.
Cf. gait with d-degree step 1→>
0/00 of motions in our model. (0 and 00
the outer poles defining d-degree 4.) It has 4 nuclei in the center
that becomes 3 in mammals.
The development of the hemispheres as 3-dimensional
volumes occurs later during evolution in mammals.
The design of cerebellum is typically 3-numbered with
3 x 2 peduncles or stalks, 3 layers in its cortex (compared with
6 in cortex of forebrain), 3 coordinate axes in structure of this
cortex, and not least its connection with the organ of equilibrium
with its 3 arches.
It's also said to have 3 representations of the
body - compared with only 2 in the forebrain.
Cortex of cerebellum differs in essential ways from cortex of
the forebrain, which has cells arranged in "radial" columns.
The big, integrating purkinje cells make up roughly
only one layer. Their dendrites become an outer layer. A big amount
of small granule cells inside the purkinje cells distribute input
to these dendrites through branching, transversal axons, i.e. axons
that become perpendicular, to the main radial input - output structure. It
correlates with the postulated circular structure that pole 4a (inward
direction) gets in d-degree 3 in the dimension model, the angle
step from 180° to 90° of polarity in relation to pole 4b.
The fact that cortex as such in its macro-shape
is much deeper furrowed than the wavy cortex of forebrain could
depend on the polarizing force from anticenter, 00 and pole 4a being
principally stronger in distal cerebellum. More of a vector
field character inwards from anticenter is retained?
We may compare the function of cerebellum for body positions, closely
connected with gravitation, and the pole 4a representing gravitation
on the physical level and in the dimension chain of forces.
There are two nuclei in the brain with
a similar cellular design as cerebellum: the dorsal cochlear nucleus
in mammals and one that receives input from lateral line organs
in fishes (Wikipedia). Hearing (and equilibrium) organs of
mammals have been regarded as developed from those lateral side
lines. These organs are both senses for pressure, connected with
gravity, the primarily inward direction of d-degree 4.
In all, as distal, mainly a sensory center, regulating and inhibiting
motor activities and as such part of the primary motor ↔ sensory polarity (see Ns
I, No. 1), cerebellum may be interpreted as an organ from
step 4a ← 3 inwards in
the dimension chain. Its system has also been described in terms
of divergence - convergence, i.e. directions Vdiv Vconv, which are
outer poles of d-degree 3 in our model.
Fig Ns-42
Fig Ns-43
(It may be added that granule cells of cerebellum make up about
3/4 of all neurons in a human brain
(Wikipedia)! Input a manifold, output unity: also a relation
between lower and higher d-degrees.
c) Mesencephalon - the midbrain:
The midbrain is small and positioned at the aqueduct between 4th
and 3rd ventricles and is sometimes seen as a part of the brainstem.
The reticular network reaches up in midbrain too.
It seems possible to see the relation midbrain - cerebellum
as neural masses that correspond to the branching of ventricles
or CSF-flows in the top of 4th ventricle:
- forward to the aqueduct to 3rd ventricle, corresponding to the
central midbrain,
- sideways to the CSF-circulation around whole brain, corresponding
to cerebellum with its mostly inhibiting function.
Cerebellum is typically "debranched"
and could in our model represent the 00-pole of d-degree 4 as debranched,
meeting "the other way around" (figure below) - and hence
developed later in evolution.
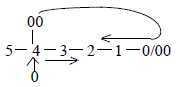 Fig
Ns-44-5-1 Fig
Ns-44-5-1
The general architecture of the human midbrain is shared with the
most ancient vertebrates. Earlier, during history of evolution,
before the craniates appeared, the mesencephalon seems to have been
the front part of the brain, the center for sight, origin for eyes
and smell organs. It's still the most front part of the brain in
birds.
It's noteworthy that the whole diencephalon and forebrain have
developed from the smell brain.
Even in lower craniates the midbrain functions
as an integration center for sensory sight and hearing, but later
these functions was taken over by centers further towards the front.
Such a fact that certain functions move forwards in the brain could
illustrate how lower d-degrees originate from polarization of higher
ones in our model.
In the midbrain motor and sensory centers become distinctly separated
areas and centered nuclei. Arrangement is the usual with motor centers
(~ pole 4b, outward direction) ventrally, sensory centers (~ pole
4 a, inward direction) distally around the aqueduct.
(We could note that number 4 here appears also
in the name of the sensory centers, the colliculus, called corpora
quadrigemina: 2 for eye movements, 2 on ventral side of aqueduct
for hearing.)
Below, in the ventral part of midbrain, the red
nuclei with functions for motor coordination appear in illustrations
as more or less circular centers, cf. 0-poles. The
difference to substantia negra should be noted:
it starts below the red nuclei and have the form of more radial,
divergent bands. This "substance" is perhaps best known
as producer of the essential transmitter dopamine. Lesion in the
function is connected with the motor disease Parkinson. Dopamine,
however, is also said to play a role in "motivation"
of species, from humans to animals as insects.
Hence, both in its shape and in function it seems
as a prolongation associated with the outward direction of d-degree
4b, with efferent vectors from the RAS system in medulla oblongata:
arousal, consciousness, potential action, with "will"
in the deeper sense (cf. above about the extrapyramidal tracts in
medulla oblongata).
The lateral axis has in midbrain been clearly marked with the doubling
of all the mentioned centers and areas, while the reticular activating
system also reaches up here. Cf. the identification of coordinate
axes in Embryology
with d-degrees 4 - 3 - 2:
4: Distal - Ventral axis: Sensory - Motor directions.
3: Front - Back: midbrain between medulla and
diencephalon: halfway separation of sensory centers (sight - hearing)
and of motor areas (red nucleus - substantia negra)?
2: Left - Right: lateral axis, doubling of structures.
Why is the lateral axis, if representing d-degree 2, expressed already
here? One aspect could be the loop version of the dimension chain
where step 4 →> 3 through
debranched degrees corresponds to step 2←1. Another that midbrain earlier was the front part
of the brain and as such included next steps too.
 Fig
Ns-45 Fig
Ns-45
Fig
Ns-46
d) Diencephalon:
Geometrically the diencephalon represent the step
where transition to 2 hemispheres occur and the step from the "circular"
3rd ventricle as a room to half-bows of 2nd ventricles. (The lateral
axis gets further expressed with the temporal lobes of the forebrain.)
Diencephalon seems to make up an inner brain in itself with the
poles 3b-3a:
- Radially, from the two symmetric thalamus structures
as centers, relaying nervous signals motor and sensory signals divergent
/ convergent to/from
the whole cortex of forebrain with neocortex.
- Circularly there is the several parts of the limbic
system above, around and below the 3rd ventricle: for instance the
bows of Hippocampus and of Fornix as a C-shaped bundle
of nerve axons from hippocampus to hypothalamus and the similar
shape of Stria terminalis. Further cortex of the cingulate
gyrus, above corpus callosum, the transverse bundle of fibers
that connect the two hemispheres.
(To this come secondary, centric bodies as Amygdala
and the Mamillary body.)
The cortex of cingulate gyrus as an inner one compared to
neocortex of the forebrain is not convoluted, and its gyri are vertical
("parasagittaly"), while gyri of neocortex are
transverse. The vertical type is observed in non-primate species
and hence regarded as older in evolution (Wikipedia).
Both these differences imply a d-degree step in
our interpretations here, from the radial - circular polarity in
step 3 - 2, to the wavy form of neocortex, implying a polarization
of d-degree 2 in convex - concave. Simultaneously it's an angle
step, here vertical to transverse planes.
One essential aspect is that here in diencephalon - as in a middle
step - the meeting of the basic nervous and endocrine systems occurs,
systems from primary animal and vegetative poles A - V:
- dorsally in epiphysis (pineal gland) which earlier in evolution
was a median eye, a photoreceptor in lampreys for instance, now
in humans is a light-dependant producer of melatonin that have with
sleep and seasonal regulation to do,
- ventrally in hypothalamus with hypophysis, which produces neurohormones
for the autonomous inner system, regulating e.g. hunger, thirst,
body temperature etc., functions of the digestive, vegetative system.
Thus, this polarity reflects primary directions
A-V: from outside environment inwards in dorsal pineal gland, from
inside the body outwards (forwards) to the ventral gland hypophysis-hypothalamus.
The fundamental coupling in these glands between
chemical and electric communication, hormones via blood system versus
nerve signals, is a polarity which can be associated with mass and
structure (d-degree 3) and covalent bonds on one hand and charge
(d-degree 2 in our model) and metal ions on the other.
The smell organ with the olfactory tract connects
here, with its enormous chemical differentiation ability.
Regarding the function of diencephalon, different
parts of it are involved in memory creation and storing.
It's noteworthy that it especially concerns memories
for spatial orientation, i.e. the 3-dimensional room, and memories
for places, in agreement with our dimensional interpretations here.
Probably sensory information from different areas
of neocortex get connected here and generally it's a well known
experience that memories need an associative "context"
to reappear. Sensations of smell do often awake memories, cf. the
olfactory tract here.
Memories as stored "inwards" could have
a parallel in the storage of DNA with methylated T-base inwards
from RNA on the genetic level.
Further, elementary emotions are located to centers
here. It's also obvious that essential emotional experiences influence
memory storing.
First such emotions are mentioned in terms of
"fight or flight" (see about
Lorenz), which can be translated to directions outwards/inwards,
the outer poles 4b - 4a of d-degree 3 in our model.
There are also centers for pleasure that activates
a repeated reward behavior, which could be described as a kind of
"circular" repetition.
Another aspect is that many emotions implies polarizations
such as openness - closeness, good - bad, negative - positive and
in this sense reflect the property of charge
as a physical quantity, in the dimension model assumed as a quality
of d-degree 2.
e) Forebrain with neocortex:
In the forebrain , newer in evolution, the polarity radial - circular
geometry appears clearly on the tissue level as a separation between
inner, radial white matter of nerve fibers and outer "circular"
surface of gray matter, the cell columns.
The surface (d-degree 2) gets wavy, meandering as described
above, (poles convex - concave and inside - outside of d-degree
2).
Transverse fibers along the surface of circumference
connect its different areas.
 Fig
Ns-47-99-2 Fig
Ns-47-99-2
The forebrain as a whole can be regarded as a circumference, a
layer around diencephalon as a central mass/space
structure. As mentioned above about cerebellum the growth of cortex
of cerebrum (telencephalon) on the embryonic stage is also
circular (Fc p. 353) as a process around a center. It has
been described as upwards on the ventral side, circular around the
front and backwards on the dorsal side, in agreement with first
directions of vegetative - animal poles when turned to a back -
front axis.
Brain parts in d-degree steps:
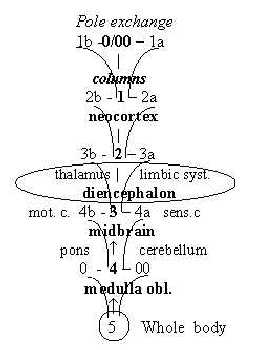 Fig
Ns-48 Fig
Ns-48
Cortex of forebrain:
The lateral furrow on top divides the primary motor and sensory
centers in accordance with the general, functional coordinate axes:
ventral side for outward direction becomes the motor area and distal
side for inward direction becomes the sensory area with visual area
at back of the head.
About numbers of things as numbers in a dimension
chain:
5 different types of nerve cells are mentioned in cerebellum, while
neocortex has a multitude: a relation few - many as between higher
and lower degrees. (According to an old classification there
were "about 50" different areas in cortex of cerebrum.)
While cerebellum (like also inner cortex of cingulate
gyrus in diencephalon as it seems) contains 3 cell layers, neocortex
contains 6 layers, as number of "potentials" in a dimensions
chain:
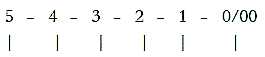 Fig
Ns-49-103-1 Fig
Ns-49-103-1
Certain of the dimensional aspects above on the nervous system
seem possible to find in the 6 layers in cell columns of neocortex;
here the layers renamed from outside inwards:
 Fig
Ns-50-103-2 Fig
Ns-50-103-2
After Nf p. 256:

Fig Ns-51-103-3
00: Unspecific sensory impulses in; general anticenter,
outermost layer.
0-4: Motor impulses = outward directed impulses out from innermost
layers.
3: Sensory impulses in to layer 3 from 3 directions: specific sensory
nerve pathways from the body, nerves from the (3-numbered) cerebellum
and from pathways along the surface of cortex from associative areas.
The cells can be regarded as interneurons and have effects on the
pathways from layers 0 and 4 according to the reference.
1-2: In loop version of the dimension model we have that debranched
degrees in higher steps outwards may meet the other way around as
steps 2←1←
00:
In layers 1-2 nerve fibers called collaterals
could be apprehended as illustration: branches from outgoing motor
axons, layer 0-4, go to layers 1-2.
Then, from layers 1 and 2 perpendicular threads
depart along the surface (note the angle step) to other columns
in cortex and have the function of lateral inhibition. Cf. interpretation
earlier of stimulation - inhibition as a polarity in step 2-1 in
the chain of all polarities
within the nervous system.
[ The order S - M in layers 3 - 2 may seem to conflict with the
view on higher d-degrees as characterized of the 0-pole and outward
direction in relation to the lower one but could be a result of
a retained polarity from step 4 - 3 in the loop model.
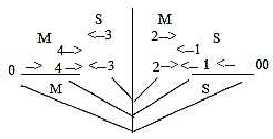 Fig
Ns-52- 104-1 ] Fig
Ns-52- 104-1 ]
Different types of sensory impulses are located to different columns.
Hence, the qualitative differentiation is radial, while the divisions
in locations are circular, i.e. which domains in the skin
the signals come from.
(According to a figure in reference Nf p. 236
one could ask if there eventually is a more fundamental division
too between a group of sensory impulses from the skin senses and
one from the inner milieu of the body, from joints, tendons and
muscles etc.?)
Conceptually the qualitative differentiation between
different senses should represent different d-degree steps in the
dimension model (cf. following files about senses).
If such identifications are possible, how are
the different qualities projected and arranged on the surface of
different columns within a certain domain? (No data in the used
references.) Consistently arranged in some way - perhaps in circles
derived from different depths of levels as in the funnel figure
here?
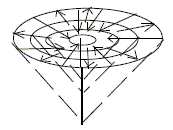 Fig
Ns-53-104-2 Fig
Ns-53-104-2
In motor cortex a certain area of columns represents direction
of movements in a joint, regulated by a group of muscles, which
get represented both vertically and horizontally (Nf p. 253).
(Perhaps in the same way as in the funnel figure above?)
A principal outline of association areas as peripheral around more
primary motor and sensory ones should with application of the funnel
figure imply that the deeper the integration center, the wider the
area for integration, the more complex the sensory impulses and
reactions. In direction upwards in the figure, along the vertical
axis, there would be more and more limited, elementary perceptions.
Cf. "tunnel vision". (?)
In bundles of nerves the nerve fibers go increasingly peripherally,
the further from the front end of the body they come from. This
arrangement agrees with the fundamental identifications here of
the front - back coordinate axis, derived from first A-V-axis animal-vegetative
poles, 00 — 0.
 Fig
Ns-54-104-3 Fig
Ns-54-104-3  Fig
Ns-55-104-4 Fig
Ns-55-104-4
Compare the patterns of growth in bird embryos around the primitive
streak and amoeboid movements through currents in the cytoplasm,
right figure above.
Psychological "faculties":
We may associate the main parts of the brain with the psychological
"faculties":
- will in the deeper sense of aim and direction with
the brainstem;
- emotions with diencephalon and the limbic system;
- conceptions of the world as 3-2-dimensional structures
and "plane" pictures with the forebrain including
parts of diencephalon and inner "cortex of cingulate gyrus";
- thoughts as linear connections between concepts
with neocortex;
- speech as thoughts transcribed into motions in last
(and every) step:
Faculties - D-degrees - Brain parts:
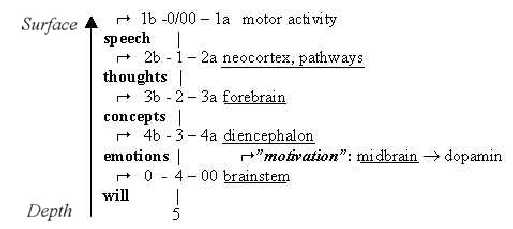
Fig-Ns-56*
Note: step 1-0/00, some kind of communication
and motor activity in each step.
Potential "speech", "thoughts"
and "concepts" should exist already in higher d-degrees and more elementary
animals without a forebrain according to the loop version of the
dimension model.
*See further a book in Psychology, "The I and the Ego" (not translated into English), connecting to these faculties and the general model. A description of the book in English here.
|