|
1.
Stepwise substantiation:
The cardiovascular system becomes
a progressively substantiated design
of the vector field of divergent
distribution in the nutrition system.
The peculiarity
that front part of intestine in
bivalves goes right through the
heart (Ez) could be taken
as a particular manifestation of
this close relation.
As
mentioned earlier a branched front
part of intestine may replace a
blood system in certain of the simplest
invertebrates.
In
simple species the circulation of
the nutrient fluid is also performed
by amoeboid cells and /or
by contractions of muscles, before
development of a blood system. Among
types of tissues (file
Levels) fluids became the
5th "tissue" at the final
step. Blood cells derive from mesenchyme
cells and reticular tissue, next
last tissue type. Later blood is
produced from bone, also a supporting
tissue of reticular type.
(The
fact that blood cells come to lose
their nucleus might be a result
of their character of last step
in a chain of tissues, anticenter
in the dimension model: their function
as only transporting vehicles?)
2. Geometry in the
development:
The development
of the blood system is a typical
example of a process through geometrically
increasing dimension degrees (d-degrees)
as in steps 0 →>
1 →>
2 →>
3 →>
4: ~ 0: scattered blood islands
from free mesenchyme cells, migrating
inwards,
~ 1: blood islands aggregating
to strings,
~ 2-3: strings developed
into tubes, where outer cells form
inner wall of the vessel (endothelium),
and inner cells become blood corpuscles.
Channels
curve in convolutions through opposite
directed currents:
~ 3: swellings
grow together into a central organ,
a heart,
~ 4: a pump with currents
outwards - inwards as a two-way
directed vector field of d-degree
4 in terms of the dimension model.
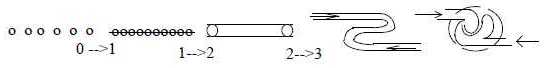 Fig
Bl-1-39-1 The transformation
of the blood canals to a heart has
perhaps (?) similarities with what
plasma physicists call "two
streams instability" (Frances
F. Chen: Introduction to Plasmaphysics,
1977): according to the author
a state difficult to analyze. The
function has a gap at vx
= 0. (So has life when the heart
stops!)
We can observe
the geometry:
- Upper, dorsal
side, representing primary 00-pole:
direction of the blood stream backwards
(~ inwards).
- Lower, ventral
side, representing primary 0-pole:
direction of blood stream forwards,
(~ outwards).
Swellings
of the S-shaped curve form through
a kind of overthrust upper and lower
heart (atria - ventricles):
 Fig
Bl-2-40-1 3. Evolution
of heart chambers:
In a simplified description the
evolution goes from a 2- to 3 -
to a 4-chamber heart, as gradual
steps of polarizations.
A
2-chamber heart is found in mollusks
and among chordates in bonefishes
(Fc p. 660).
The
evolution among chordates goes from
a tube-shaped heart in lancelets
and proceeds to a 3-chamber heart
in amphibians, a "3.5"-chamber
heart in most reptiles (2 atria
and a not quite divided ventricle)
and to the 4-chamber heart in birds
and mammalians with complete division
between the arterial blood in the
left half, venous blood in the right
half. (Biologists may argue
that a 4-chamber heart was triggered
by life on land requiring lungs
and with that a double circulation,
but one could imagine the reverse:
that dimensionally given polarization
steps led to both lungs and the
subsequent double circulation, which
allowed the general upward direction
from sea to life on land and in
air. Why didn't animals stay in
the water?! )
4.
Heart in humans:
Divisions
of the 4-chamber heart agree with
the polarizations in coordinate
axes of the embryo: Front - Back
(from Animal - Vegetative poles)
and Left - Right.
Two
divisions give 4 parts, the atria
and ventricles, and diagonally as
in 45° in complex combination
the big systemic circuit and the
small pulmonary circuit.

Fig Bl-3-41-2 There
are notable details that seem derived
from those primary poles.
The
coordinate axes Front - Back as
derived from A-V-poles represents
a secondary axis 00 <=====>
0 (file Embryology.
This implies in terms of the dimension
model that we get
- outward
direction from the ventricles
that have the lower, rear position,
corresponding to original vegetative
0-pole, with blood forced forwards
towards the 00-pole,
- inward
direction from atria
that have the upper, front position,
corresponding to original animal
00-pole, with blood forced downwards,
backwards towards the center pole.
This in spite
of the fact that it entangles the
connection to blood canals. We
have also that it is the ventricles
(from 0-pole) that during evolution
get divided later. (Polarizing force
from anticenter in our model.) The
very triangular form of the heart
as a whole corresponds to a 0-00-polarity
with apex as center closer the back
pole, the breadth at the front.
Apex is also turned more to the
ventral side.
The
heart as a 3-dimensional organ,
polarized mass (muscles) —
space (with blood), shows also the
radial structure (pole 3b in the
dimension model) in its papillary
muscles that depart from near bottom
of ventricles (the apex). Another
detail is that the right ventricle
is partly bent around the left one
(Mf p.105), which seems guided
by the venous direction inwards,
hence reflecting the function of
00-pole as anticenter and inward
direction in relation to the 0-pole
as center and outward direction.
A similar
feature is that atria from inward
direction have thinner walls (highly
expandable) and can be more flattened
(Aph) - as d-degree 2 in
relation to ventricles of d-degree
3 with the aspects here.
Still another, similar feature is
that the channels for flows outwards
from both ventricles are centered
at the median of the heart, while
the inflows are peripherally located:
also a feature of center - anticenter
relation. In addition, the
fact that inflows (have to) pass
an antechamber before entering ventricles,
while the outflows go directly to
canals, can be interpreted as an
illustration of origin of directions
in a dimension chain:
- Inflows
via atria as 3-dimensional rooms,
i.e. from anticenter and lower d-degree
towards d-degree 4.
- Outflows
from higher d-degree 5 and 0-pole
to d-degree 4 and straight to canals
that can be regarded as substantiated
structures of d-degree 4b.
Valves:
The aortic
and pulmonary valves for outflows
are both 3-lobated, the half-moon
shaped cusps.
The
valves for inflows however differ
in a way that seems to reflect the
opposition between the big systemic
circuit and the small, pulmonary
one. It's
3-divided
in tricuspid valve for venous blood
from the big body circuit,
2-divided
in bicuspid valve for the small
circuit with blood from lungs.
It happens to correspond also in
number with our interpretation of
the coordinate axes (file Embryology)
as representing d-degree 3, Font
- Back, and d-degree 2, R - L respectively.
It's surely interpreted
in other, more physical terms by
biologists. Yet, there is the similar
3-2-relation between lungs: right
lung with 3 lobes, left lung with
2 lobes. (A division of number 5!)
Hardly referable to the same physical
causes. Neither could the fact that
2/3 of
the heart is located on the left
side of the middle be an explanation.
Why this asymmetry? It indicates
sooner that asymmetries appearing
along the coordinate axis L - R
have a deeper root, inherited from
original complementary poles of
higher d-degrees in terms of the
dimension model. There is the asymmetry
too in ways of arteries to head
and body (as there is between the
cerebral hemispheres).
5. Canals - the vascular system:
The big, systemic circuit
is mainly branched 180° along
the F-B-axis (front - back). The
small pulmonary small circuit to
the paired lungs branches naturally
along the R-L-axis (right - left).
With the
earlier
view on these axes as representing
d-degrees 3 - 2 it could be noted
that it's connected with the d-degree
step in
phases: between exchange
of chemical molecules in the big
circuit and of gases (CO2
- O2) in
the small circuit.
Number
relation 2 - 1:
Arteries
and the deep veins are together
enclosed in a capsule of connective
tissue as an expression for the
two-way Direction of d-degree 4.
Often 2 veins
go parallelly with the one artery,
a number relation 2 to 1 between
inward direction from 00-pole and
outward direction from 0-pole. These
data concerns humans.
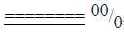 Fig
Bl-4-42-2 Fig
Bl-4-42-2
Similar 2-1
relations appear in the canals of
lancelets, the simplest chordate
with a developed blood vessel system:
Ventral canals,
both front and back parts, are single,
unpaired canals.
Dorsal
veins are paired. So is front part
of dorsal artery canal, not its
back part (resembles a fork).
Dorsal side represent
primary anticenter, the animal 00-pole,
the front part the secondary anticenter
(00') in the embryological development.
Hence, the doubling of canals (veins)
or branching of canals (arteries)
seems guided by the 00-pole, in
opposition to ventral 0-pole representing
singularity. The opposition 2 -
1 affects both axes: Distal - Ventral
and - in distal artery - the Front
- Back axis. (A simplified figure
below after Kz p. 19.)
In the dimension
model the 00-pole, from which follows
inward direction, is defined as
primary polarizing force. It's difficult
to imagine any biological reason
or other necessity for the duplication
in inward direction (distal, front,
blood direction in veins), than
the simple numerical one: the polarization
of 1 to 2. 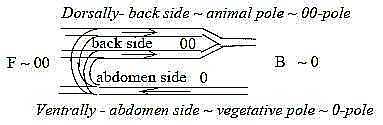 Fig
Bl-5-43-1 Fig
Bl-5-43-1 A radial
- circular polarity:
In the human body there is a net
of superficial skin veins without
corresponding arteries, (Mf p.
117) as a kind of "circular
structure" of d-degree 3 towards
the surface, while the arterioles
instead are branched more "radially"
outwards: a polarization of type
3a-/-3b
in the dimension model.
Dimensional
steps in size of arteries:
With biologists' designations we
get a whole chain of dimensional
decrease in size, (simultaneously
as the ramification of vessels increases
the dimensional structure as a whole
1→
2 →
3):
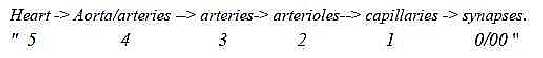 Fig
Bl-6-43-2 Simplified,
4 types of tissue layers can be
distinguished in the arterial system:
5-4: Heart: striated, special musculature
of branched cells.
4-3: Aorta
- thicker arteries: 3 layers with
intermediate layers of mostly elastic
threads.
3-2: Thinner arteries
and arterioles: 3 layers, intermediate
layers of smooth muscles.
2-1:
Capillaries: 2 to 1 layers, outer
layer only a net of reticular web.
1-0/00:
Synapses: Transportation through
walls of capillaries to/from
the tissue fluid.
The
"pole exchange" outward/inwards)
through the walls of the capillaries
from the arterial to the venous
system implies a kind of reverse
relations in hydrostatic and osmotic
pressure between arterial and venous
capillaries (Zf), which could
be regarded as expression for a
"pole exchange" in terms
of the dimension model in last d-degree
0/00 of
motions.
(The surplus of outflow pressure
is taken care of by the evolutionary
later developed lymphatic system.) From
vibration to rotation:
It
is noteworthy that the blood flow
at an early stage of evolution (e.g.
in annelids) is bi-directional:
one moment inwards, next moment
outwards in the same vessel à
la vibration. It develops during
evolution to the circular system
with separated out-/inflow
vessels, what may be apprehended
as a rotation, a 2-dimensional motion.
 Fig
Bl-7-41-1 Fig
Bl-7-41-1 The development
corresponds to the presumed d-degrees
of motions
in d-degree 4 and 3 in our model.
We get outward - inward flows, poles
4b - 4a (from 0- and 00-poles respectively)
of d-degree 3 in different canals.
The appearance
of special lymphatic vessels, later
in evolution, implies a secondary
polarity of the character 00-0,
here between veins and lymphatic
vessels: the opposition blood
from cells as centers in veins
versus blood from extracellular
fluid (anticenter) in the lymphatic
vessels. Vibration →>
Rotation →>
Translation in 3 dimensions?
A 3-dimensional motion as "translation
in 3 directions" - could perhaps
be identified with the further branching
of vessels in the whole body - and/or
more specifically the capillary
networks in all tissues as the 3-dimensional
motion presumed in d-degree 2.
The lymph:
With the evolution of a lymphatic
system, we have once again the relation
2 to 1, here in number of systems:
2 systems for inward direction,
1 for the outward direction. The
lymphatic system connects to the
venous system.
In
our model the 00- pole represent
the primary polarizing force, upholding
potentials when in balance with
the integrating 0-pole or when stronger
breaking them.
This
property could be seen expressed
in the role of lymph in the immune
system with activities of macrophages
etceteras. Geometry of lymphatic
nodes seems to reflect proposals
in the dimension chain:
The
nodes have one convex side, one
concave, which is one of the geometrical
polarities of d-degree 2 proposed
in our model: 2a convex →>
2b concave.
In
agreement with this geometry vessels
inwards the nodes go to the convex
side while the outgoing vessels
depart from the concave side.
Further, there
is the polarity of many incoming
vessels, few outgoing ones as secondary
manifestations of the 00- and 0-poles,
a-poles versus b-poles. Pathways
of lymphatic vessels show the same
bilateral asymmetry as arteries,
in fact a rather curious asymmetry:
right side vessels come mostly from
right side front, head and arm,
while left side vessels comes from
the whole body, trunk, intestines
and head and arm on left side. There
is a certain similarity with ventricles
of heart: right ventricle pumping
blood to lungs, i.e. only a front
part, while left ventricle pumps
it to the whole body. Yet, this
cannot explain the asymmetry of
pathways out in the body.
(If
left hemisphere of the brain governs
muscles in vessels of right side,
shall we then assume that it doesn't
care about the whole and only manages
to serve half of the front?)
6. The liver:
The
liver develops during evolution
from a tube-shaped gland to a separated
3-dimensional organ. It could be
said to represent the very transition
between the nutrition and blood
systems. It continues the breaking-down
process of nutrients in the alimentary
canal (proteins, lipids) but performs
also synthesis, for instance of
a carbohydrate as glucose. (Hence,
backwards relative the process of
glycolysis.)
As
the liver has a double-directed
performance of breaking-down and
synthesis, it has double exits:
excretion of dross products via
the bile in one direction and distribution
of nutrients in the other direction,
through vena cava.
It
is regarded a part of the venous
system but blood from both arteries
and veins enter the liver and merge
in sinusoids: roughly 1/3
and 2/3
respectively (Aph, p. 890);
note the returning 2-1-relation
if so. According to other sources
the quotient is circa 1/4
and 3/4
(Mf, Wikipedia). Geometry
of the liver shows up to be remarkably
regular and strict internally:
It is the most
massive gland and its cell masses
and blood rooms, the sinusoids,
can illustrate the polarization
mass - space of d-degree 3 in our
model.
Each
lobule has a hexagonal (5 - 7 edges)
shape and the blood canals and fluid
directions illustrate to an exceptional
degree the fundamentals of step
4 →>
3 in the dimension model: inward
direction from anticenter, that's
from the corners in the hexagons,
outward direction from the center,
the central vena cava inferior.
There is simultaneously the
polarity between a manifold inwards
from anticenter versus unity from
the center.
The
lobules illustrate further the geometrical
poles of d-degree step 3-2 (3b -
3a) in the radial arrangement of
cells versus the circular blood
rooms. 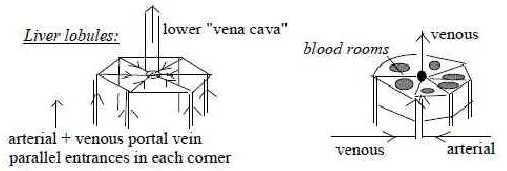 Fig
Bl-8-45-1 Lobules are
plates, only 1 cell thick, i.e.
2-dimensional. Thus, they give a
picture of how radial/circular
poles 3b-3a could characterize d-degree
2 in a way not presumed before in
the dimension model. In their 3-dimensional
storing they show at the same time
the polarities of higher d-degrees
4 and 3, center-anticenter, outward-inward
directions. (On the macro-scale
the liver is divided in 4 lobes
of different sizes, right - left
- quadratic and caudate lobes, as
if mass was differentiated along
a separate dimensional chain 4-3-2-1.
With size associated with d-degrees
we could imagine two levels: a)
d-degree 5 polarized 4-1= quadratic
+ caudate lobes, b) 5 polarized
3-2 = right and left lobes.
It's said (Aph
p. 891) that each lobe contains
about 105 lobules, in
number of 10-powers as from a dimension
chain.) Cells in the liver
can have several nuclei, what is
called plasmodia, the fission type
where nuclei divide without division
of cell plasma; this in opposition
to the multi-nucleate muscle cells
of the heart that are the result
of fusion between individual cells,
syncytium (Kz p.150).
This polarity
fusion - fission on the cell level
agrees in directions with the relative
polarity F -B (00-0) of mesodermal
muscles from front somites versus
liver as a gland from vegetative
pole. The same polarity is expressed
in positions of heart versus liver,
on opposite sides of diaphragm.
It's notable that
liver cells also in human beings
have the capacity to regenerate,
showing a highdimensional potential.
7. Liver and lungs:
Gills
and lungs belong naturally to the
blood system. Liver and lungs illustrate
in several respects the differences
between higher and lower d-degrees
and the complementary polarities
derived from 0- and 00-poles in
origin, directions, positions and
shapes:
- Lungs develop
from front part of the alimentary
canal, partly from ectoderm,
the liver as a gland
from central part of endoderm. -
Lungs are positioned in front part
of the body, liver in back part,
below diaphragm.
- Lungs
are pairs, liver an unpaired organ.
- Ways of the blood: in lungs
one-way directed venous →>
arterial, v →>
a,
in the liver
v + a →
a plus the double direction to bile
and vena cava inferior.
- Phases of substances: in
lungs exchange of gases, in the
liver fluids and organic molecules.
Phases:
 Fig
Bl-9-46 Fig
Bl-9-46 Geometrical arrangement
of blood vessels can illustrate
the complementary polarities of
d-degree steps 2-1 versus 3-2 in
the dimension chain: In lungs as
an organ of the surface blood vessels
get structured as half-spherical
nets outside of and around the alveoli*,
with gas exchange between outside
and inside as poles 2a - 2b. In
the liver the vessels are organized
vertically and radially along perpendicular
coordinate axes with blood in the
inner "rooms", both features
of poles 3a - 3b.
*Compare the kidneys where the blood
vessels themselves get shaped as
balls within a bowl-shaped capsule,
a kind of inversion of the structure
in the lung alveoli. Number
5 again:
In the lungs
the number 5 appears again as so
often in biology- and the asymmetry
left-right. Together the lungs have
5 lobes, divided 3 (right lung)
- 2 (left lung). (Right bronchial
tube goes also more straight downwards
as a direct continuation of the
windpipe.)
Each
lung is then divided in 10 segments
with an own bronchus to each, divided
on the 3 right lobes 5 - 3 - 2 and
on the 2 left lobes 5 - 5. (fusing
to 8).
To 06. Muscles
|