|
1. Some general aspects:
1. A cell is a complex center, a "0-pole" in relation
to its environment. At the same time it is a whole in itself, as
dimension degree (d-degree) 5, ~ 0/00
in the background model
applied here - including all the geometrical polarities of the dimension
chain.
Chemical substances could be regarded as both
building workers and building blocks that substantiate and illustrate
an underlying drawing.
The enormous complexity in a cell seems only possible
to understand if we regard it as results of internal polarizations
within some already enclosed unit with a substantiated anticenter,
an original structure of type 0/00, center/anticenter
(c/ac); the complexity a result of opposite, internal forces.
(Which kind of center, which kind of anticenter
when first cell appeared?)
2. Why a relation between DNA or RNA and proteins as a genetic
code?
It's one main question. Here the c - ac relation between
C-atoms in tetrahedrons of amino acids (ams) versus rings of the
codon bases are stressed.
It is suggested that the relation can be illustrated
with the hexagonal pattern below. Imagine a layer of graphite, or
- with virtual free valences upwards next layer - a crashed diamond.
(All substituents to C-atoms dismissed.)
Carbon rings, sharing edges, and centers with
3 radii (+ the virtual 4th) become two aspects on the same whole
pattern. Two ways of reading.
(Other main classes of substances sketched as
well.)
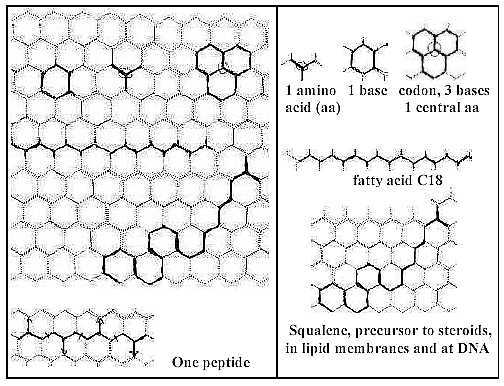
Fig C-1-7-Hex
One center with 3 radii is defined by 3 rings - as
amino acids by codons.
The horizontal peptide in the figure above can
illustrate how side chains of amino acids must point in opposite
directions for each amino acid to get a separate triplet of bases.
Codons too get opposite directions:
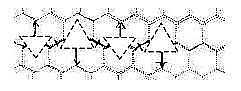
Fig
C-8-Hex
(Eventual implication for interpretation
of proteins?)
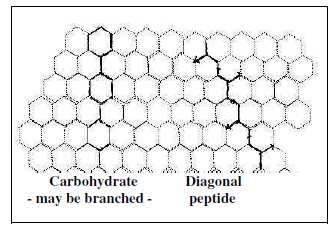
Fig C-9-Hex
Amino acids (3 of them) give main contributions in
synthesis
of the bases. The simplest amino acid Gly makes up the very
center at synthesis of the purine bases A and G. Asp makes up about
half the pyrimidine bases. (Moreover, an amino acid as Ile is used
in synthesis of fatty acids.)
One 6-ring in the bases gives theoretically,
2 x N-C-C, the bound backbone chain in an amino acid. (See further
The protein
synthesis.)
Mean mass value of a codon base (RNA) is 126
+1, 6 x 21 +1. Mean value of a side chain of amino acid is approximately
half this number, 63, 3 x 21. (About "A-Z"-numbers
here.).
3 units with a central C-atom and 3 radii shape
a 6-ring. Cf. about mass number transformations in the
genetic code where 8 bases give the backbone chains or
side chains of 24 amino acids, a relation 1/3.
RNA-chains have been found to possess a certain catalytic capacity,
which may be easier to understand with the hexagonal illustration.
In our cells it's the halving of 6C carbohydrates
(however transformed to 5C rings + 1C, fructose) that through glycolysis
and citrate cycle leads to synthesis of fatty acids and amino acids
respectively.
About fatty acids, the illustration can be compared
with the fact that the double layer of lipids in cell membranes
at a certain temperature (~ 37°) get hexagonal structures, an
unstable state of phase transitions (Zf).
The figure doesn't include pentagon rings, the extra 3 edges in
purine bases, not differences in angles of valences, nor any substituents.
With the figure above it's not the intention to assert that this
was 'the way it happened'. Yet, it may illustrate the close relations
between substances and the underlying two aspects on the whole structure:
a pattern of rings or a pattern of centers with 3 radii.
The idea of a crashed diamond leads to the thought of different
substances created from the different ways and diffraction of light
in a crystal.
A speculation about phase waves - a very
abstract thought:
From a hypothetical reasoning in a
file on physics it could perhaps be possible to apprehend
proteins (and other classes of substances) as a kind of "phase
waves" on a superposed, substantiated level: expressions
for relations between other substances as "waves" .
Or DNA (RNA) as materialized phase waves between protein chains?
(Cf. about the Balmer series for spectral
lines of hydrogen, fig 1-1.) It contradicts of course
scientists' opinion that such waves don't contain any information.
(If the mass numbers of codon bases could be
associated with d-degrees or steps between d-degrees in different
ways, see the
Genetic code, and these steps also include angle changes,
then the relation between the nucleotides as "rotating vectors"
could give birth to phase waves? Such waves - or complex relations
between them, could be imagined to decide the row of amino acids
in a protein - or follow from them?)
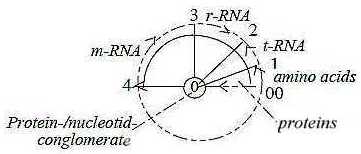
3. The protein synthesis:
Compared with the view in the hexagonal pattern above and the very
close c/ac-relation amino acids/bases,
the synthesis
of proteins in a developed cell appears as a much more
complicated process "the other way around":
 Fig
C-10 Fig
C-10
Illustrated according to the loop version of the model
- or as in a circle:
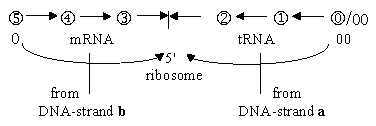
Fig
C-12
C-11-144-1
One theory among scientists about origin of the genetic
code assumes a preceding stage of close connection between codons
and amino acids before the transcription system developed. In secondary
forms a close relation appears in DNA rolled up on histones, in
ribosomes and in tRNAs, binding individual amino acids etc. Could
perhaps the existence of the special membrane-enclosed nucleol
within the very nucleus of the cell, at a certain chromosome,
containing material for the ribosomes (rRNA), reveal something about
such an underlying origin of closer dependence?
Why this complicated, circumventing process to produce proteins?
While amino acids directly engage in constructing bases, contributing
most of them, the way from bases (ac-pole ~ 00) to amino acids (c-pole
~ 0) becomes very different, a "code", as a reference
over the distances created by the separation.
One aspect with reference to the dimension model
could be that outward direction from a 0-pole (c →>ac) implies radial structure in d-degree 3,
while inward direction from a 00-pole appears as 'circular'.
These complementary geometric forms could have decided the geometry
of the very processes (?).
4. A cell as inversion of an atom:
The cell as the "elementary particle" of biology can
be interpreted as an inversion of an atom. Its outer cell membrane
may be seen as a potential barrier, an inversion of the nuclear,
strong force (FST).
- The lipid bilayer membranes keep together solely by hydrophobic
bonds, an H+-aggregating, gluey force, while proteins,
DNA and other inner structures are built by the covalent force
between atoms with unsaturated electron (e-) shells.
(About views on chemical forces here.)
H+ and e- become elementary
"carriers of forces" on the chemical level.
- Charges are reversed, with relative negative charge on
inside, positive on outside.
(Positive (+)-charge seen as originating from inward direction,
negative (-)-charge from outward direction.)
A cell membrane as a surface is "semipermeable". It's
characterized by changing phases or degrees of permeability. Permeability
as a physical quantity is inversely proportional to charge squared.
CF. d-degree 2 as surfaces and charge
in this model assumed as a 2-dimensional physical quantity.
- A cell in its main structure shows also a reversed relation mass
- space in the relative sense of phases: membranes as anticenter
of the cell around inner space. In atoms the protons (H+)
represent most of the mass, while outer electron shells make up
most of the atomic space.
About other suggested inversions, see file
1/7, and mass number 544 in the
genetic code as from the inverted p/e
quotient.
(All physical
forces, which in these files are interpreted as developed
out of one another, must reasonably be expected to appear in the
cell in their chemical versions. And naturally, a cell can be
described in all physical quantities
interpreted as steps in a dimension chain. (Density introduced
as a first such quantity defined by the polarization center -
anticenter, in later step expressed as mass per volume, in step
1→>0/00
as distance per time and frequencies.
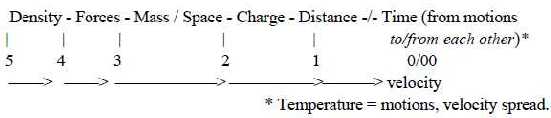
Fig 13-145-1 )
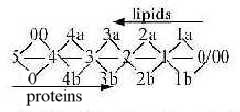
5. Proteins - Lipids, the macro-level structure:
In the cell, on its macro-level, the proteins come to form the
main radial structure, the cytoskeleton, while the fatty acids as
lipids make up the roughly circular structures, the primary cell
membrane ( in eukaryotic cells the nuclear membrane, the endoplasmic
reticulum (ER), Golgi apparatus etc.)
It's the two main structure-building parts
of the cell and the geometrical polarity radial - circular
as poles out of d-degree 3 in the dimension chain.
(It's not the whole truth, since protein threads
are also found as "horizontal" layers inside lipid membranes
an as nets outside these on some kind of cells. Yet a main polarity.)
 Fig
14-143-1 Fig
14-143-1
Protein threads as F-actin and microtubuli stretches radially outwards
through cytoplasm and likewise in cilia and microvilli. Other proteins
form the radial canals in lipid membranes. They function as infrastructure
and transport tracks, as for instance spindles at cell division
obviously also as vectors.
At protein synthesis the peptide chains grow 'straight
through the walls' of lipid membrane
in the endoplasmic reticulum in a way that feels curious, if not
founded in a basic geometry.
Radial structures are principally unlimited outwards, circular
structures closed. It's the same polarity as between electric
and magnetic field lines E and M. Cf. in file
about forces
relations to protons (p) and electrons (e) within plasma physics:
p ~ M2
e ~ E2
Hence,we find a connection between magnetic fields squared, protons
(H+) and the closed H-bonded lipid membranes of a cell,
as between electric fields squared, electrons (e-)and
covalent bonds of proteins.
The fact that the cell membrane in itself is
a structure of both kinds, containing about 50 % proteins, 50 %
lipids, could remind of light beams with their regular phase
displaced changes between E- and M-factors. Compare again possible
readings in figure 1 above.
In the dimension model the radial and circular poles of d-degree
3 may be seen as derived from 0- and 00-poles respectively in a
haploid chain, illustrate in the figure below, however somewhat
misleading: the steps for lipids sooner should be angled more perpendicular
to the steps for proteins.
Amino acids in protein derive from molecules of
5 - 4 - 3 C (carbon), while the synthesis of fatty acids is a repetitive
process in step 3 - 2: (3 C -1 C→>
2 C) to n x 2 C. (Cf. 2 as d-degree of a border, a surface.) It's
a synthesis on a multienzyme complex with amino proteins involved
and amino acid Cys providing the HS-sites for attachment during
the process..(Cf. Cys, mass of side chain 47 A, total mass of side
chains of amino acids from 24 differentiating codons = 32 x 47.)
 Fig
C-15 Fig
C-15
In geometrical macro-structure the lipids develop in steps 1 -
2 - (3): from linear fatty acids, d-degree 1, to 2-dimensional when
with backbone chain of glycerol (3 C) to the spherical membrane
enclosing 3-dimensional volumes, demarcating different phases and
rooms.
About glycerol and polarization of carbohydrates,
6 C, into 2 x 3 C, see below.
An essential process as from middle step 3 - 2 is the Pentose-phosphate
cycle, that implies transformation of 5 x 3C into 3 x 5 C (note
number 15, sum of d-degrees in the dimension chain), where one way
is 6C - 2C to 4C, + 3C to 7C, - 2C. (7 = sum of poles in d-degrees
3 an 2.)
It can be added that when plants build in 1C from
the air in a 5 C carbohydrate to 6C, it is in the middle between
3rd and 2nd C-atom.
[About classes of substances on the molecular level
here.]
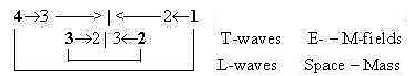
6. L-waves, T-waves and subunits:
Fatty acids as subunits of lipids have a similarity with longitudinal
L-waves, although in a zigzag shape. (Cf. perhaps the movements
of single P-lipids along the membrane during its fluid phase.) Protein
chains, unfolded, with their side chains of amino acids, correspond
in a similar way to transversal T-waves.
It's presumed in the model that L-waves as linear,
1-dimensional, [→ ← → ← → ← ...] are connected
with forces FG and FA,
gravitation and the complementary outward acceleration force. These
represent d-degree 4 in a dimension
chain of forces and are interpreted as polarized into positive
and negative (rather antipositive) curvature in step 4
→> 3 in the model
here and appear in the polarity Mass and Space in terms of physical
properties.
Membranes are connected with the polarity Mass
- Space in the secondary sense of differentiating phases. Their
H+-bonds (methyl bonds), connected with mass, are identified
as pole 4a among chemical
forces, i.e. inward direction. The bilayers, however, of
fatty acids in membranes can in fact be seen as illustrating both
the inward and outward directions of d-degree 4.
(Cf. about linear waves as motions in section
8 below.).)
T-waves are exemplified by the more well-known
electromagnetic waves and the force FEM,
[↓ → ↑ ← ↓ → ↑ ← ↓
→↑...] with the complementary electric and magnetic factors FE
and FM. It's presumed as a force developed
in following step 3 - 2 in the model.
The covalent bonds in proteins are identified
as pole 4b in the dimension chain of chemical forces, implying outward
direction. Peptide bonds include polarity of charge (NH3+
↔ COO-) and represent the L-factor in proteins
as T-waves.
With these identifications the lipids come to represent a deeper,
underlying level in relation to the proteins, where we for instance
also have an angle step in the phase displacement between field
components from 180° to 90°.
How to regard this contradiction? The only way
(?) seems to be to regard debranched degrees from higher steps meeting
"the other way around" in accordance with the loop model
.
Fig
C-16
As waves proteins and lipids should correspond to a substantiation
of the dimension chain of motions:
- Step 5 → 4, physical
quantity Density proposed, 1 degree debranched, linear waves as
polarizations of density. (Bilayer of P-lipids as micells appear
at higher density.)
- Step 4 → 3, Direction,
d-degree 4, polarized to a perpendicular angle , 90°: T-waves,
2-3-dimensional depending on aspect.
Both wave types contain the features of both poles of the higher
d-degrees→ 4 →
3. Backbone chains of the proteins (typo L-waves but expressed in
quantity charge, d-degree 2) become expression for divergent inner
space (pole 4b), sustaining it as radial cytoskeleton.
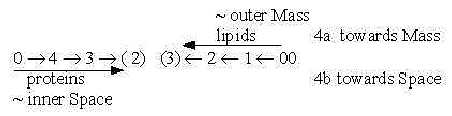
Fig
C-17
Waves as such could probably be best
understood as the sewing together of the two complementary fields
in each d-degree.
If the aspects above on lipids and proteins as corresponding to
the polarity between L- and T-waves, it leads naturally to other
speculations. See section 8 below..
(It has to be added here that Archae bacteria, assumed as the oldest
unicellular phyla, uses isoprenes, molecules of 5C, instead of fatty
acids of n x 2C in their membranes. It implies carbon groups as
a kind of transversal factors in their lipids too.)
7. Polarizations into complementary "poles"?
According to the dimension model a developed cell should be possible
to interpret as the result of polarizations into complementary "poles"
as partial structures in different -degrees and on a lot of levels.
When it concerns the elementary definitions: opposite vector directions,
mass - space, charges +/-, outside - inside,
divergent - convergent motions etceteras, all kinds of complementary
polarities can too easily be found in a cell - but how about classes
of substances on a more concrete level?
Carbohydrate - fructose:
The most obvious example is the division of carbohydrates: the halving
of an hexose (fructose), leading to lipids and amino acids of proteins,
i.e., the main circular versus radial structures of the cell.
Two 3C pyruvate +/-1
C gives
a) C4 (note sign +, inwards) to mitochondria to oxaloacetate
in citrate cycle (note a cycle), and
b) outwards (sign -) C2, Acetyl(~Coa,) leading to to fatty
acids and isoprenes.
 Fig
C-18 Fig
C-18
In this process the glycolysis also gives the polarity
between perpendicular parts of triglycerides (glycerol versus fatty
acids).
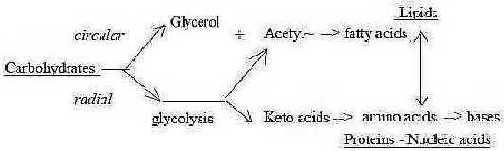
Fig C-19-147-1
In a secondary or synthesizing way we find the branching from Acetyl(~CoA)
to elements in the two classes fatty acids, (the repeated process
C3 - C1 to n x C2), and isoprenes/steroids
(3 x C2 - C1 to n x C5). The amino acids Ileu and Leu that only
differ stereometrically, contribute to fatty acids and steroids
respectively.
(The ways for Acetyl(~CoA) has also a preceding
ramification, where one joins the citrate circle, the "protein
way", leading back to 6C - 5C molecules, here as keto acids.)
DNA-strings:
The division of a carbohydrate as fructose leads to different
classes of substances. When it concerns internal polarizations within
the classes, DNA seems to be the only (?) or most obvious structure
that corresponds to an unpolarized "d-degree".
It's opposite strands represent codons of mRNA
and tRNA gets polarized into complementary units or "poles";
meeting "the other way around" on ribosomes as mRNA →>
← tRNAs at the
protein synthesis.
The opposite directions of strands lengthwise
and the complementary forms of the bases underline the polarities
as does the complementary syntheses
of the bases.
Complementary directions in structures:
Within classes the most obvious examples of opposite directions
in structures are DNA strands and bilayer of lipid membranes between
inner and outer rows of P-lipids.
There are striking similarities between these
two molecular compounds with very different functions. Both consist
of a) P-group, b) a carbohydrate element, c) then the specialized
molecular objects: codon bases and fatty acids respectively:
P-group →
deoxiribose → base —>
H-bridge <— base ← deoxiribose ←
P-group
P-group → glycerol →
fatty acids —>
H-bonds <— fatty
acids ← glycerol ←
P-group
The POxHx-groups surround both structures as "first and last"
elements - as do the poles 0 and 00 out of d-degree 5 in the dimension
model.
Notice P-atom, valence 5. and d-degree 5, polarized
0 - 00, origins for primary binding and polarizing forces in the
model. (Cf. too fructose-6-phosphate that first when enclosed between
two P-groups in fructose 1,6-bisphosphate can be halved by an enzyme.)
Another similarity may be seen in the perpendicular,
vertical - horizontal bonds between P-groups and the carbohydrate
part.
Simultaneously there is the principal difference
between "linear" structure of glycerol and fatty acids
in lipids and the ring closed corresponding parts in DNA, like a
step from d-degree 1 to 2. And ribose in DNA, 5C, presupposes the
pentose-phosphate cycle 5 C3 →>
3 C5.
About the different parts, it seems that the complementary relation
between inner bases of the DNA-structure above could have correspondences
in the outer, different kinds of attached molecules to "heads"
of P-lipids on inner and outer layers respectively of membranes
(Wikipedia, Lipid_bilayer).
We can also remember how the chains of fatty acids of lipids arrange
in hexagonal patterns - like the one in codon bases - during certain
phases. Geometrically it appears as a d-degree step 1 to 2.
The correspondence between these central versus anticentral constructions
needs certainly a better interpretation.
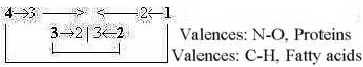
One view in terms of valences, to compare with
the figure above on lipids and proteins as L- and T-waves. C - H
valences 4 --- 1, N - O valences 3 - 2:
 Fig
C-20 Fig
C-20
Polarities within lipids and proteins:
Compared with DNA a similar structural division of lipid
membranes seems not to exist (?). If there is any complementary
features between P-lipids of inner versus outer layer, in origin
or construction, it doesn't seem recognized yet. There is however
other kinds of internal polarities:
One is the transition between phases, which both can
exist simultaneously on different parts of the membrane: the solid
gel phase and the fluid one - as if area of the membrane had inherent
subdivisions (?).
Another opposition is the just mentioned changes between
ordinary and hexagonal structures (Zf p. 181).
A third feature, more adequately identified as a polarity, is the
ability of lipid layers to form "plain and purl" structures,
heads outwards or inwards: the polarity of directions out of d-degree
4. It's also a polarity of charge out of d-degree 2 in the model.
The geometrical polarity of d-degree 2 has been defined elementary
as outside/inside. On a macro-scale
the bilayer membrane illustrates it. The sense or function, if any,
of such inversions seems not clear.
One older observation was that fragmented membranes
of mitochondria appeared some with "knobs" on the outside,
some with these knobs on the inside, thus examples of such polarities
and inversions.
Evaginating/invaginating parts
of the membrane forming vesicles illustrate one kind of 3-dimensional
motion, assumed in structures of d-degree 2 and at the same time
the polarization of directions of d-degree 4. Cf. meteorology: high
and low pressure cells debranched from Rossby waves around the North
Pole.
Among proteins, there are one main polarity between synthesizing
and breaking enzymes.
Another fundamental one seems to be the one between
proteins as transporting vehicles, some transporting molecules outwards,
some inwards - as kinesin and dynein in cilia tubuli. They illustrate
the opposite vectors of d-degree 4. (It would be very interesting
to know which factors, amino acids or codons or other factors in
such specialized proteins that are decisive for their complementarity.)
Certainly many other such examples of complementary
proteins exist, also acting on long distances from one another.
Are such complementary pairs of proteins eventually a fundamental
principle rooted in the genes?
Human genome includes more than 100,000 proteins,
while number of genes is only about 28,000 according to some data.
It means that one gene on average should code for about 4 proteins.
Through different frames at transcription or different cuttings
of mRNA before? Do proteins from the same gene eventually show on
polar relations in their functions?
It's hard to imagine any common x-dimensional
unity as origin for complementary proteins, if not in opposite pieces
of DNA-strands or perhaps in complementary folding?
(About classes of substances in cells, see further Biochemistry.).
8. Speculations:
About L- and T-waves:
If we accept the hypothetical view in No. 6 above on fatty acids
and proteins as substantiated L- and T-waves, and these are taken
as connected with the pairs of forces
FG - FA and FE
- FM respectively, it awakes two kind of
questions.
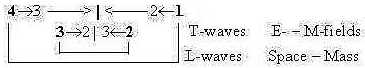 Fig C-21
Fig C-21
 Fig
C-22 Fig
C-22
A first question is if proteins in any
way or sense could be seen as derived from or later than lipids
(!) as lower d-degrees from higher ones? Nothing in actual, internal
biochemistry of a cell supports such a view. As atoms N and O derive
from C in the carbon-nitrogen cycle of fusion in the sun but that's
another history. Could some first amino acids have been constructed
on, in or between bilayers of P-lipids? We can only point to some
close relations: the attachment of ribosomes to membranes when protein
synthesis occurs, the protein chains growing straight in through
the membranes of the endoplasmic reticulum, processed further within
the membrane enclosed rooms, as in the Golgi apparatus - and the
~50 % proteins curiously 'interrupting' the outer cell membrane
itself.
The second question is if expressions for the pairs
of connected forces in some way can be identified in the lipids
and proteins.
About lipids, we could ask if the inner
layer of a bilayer membrane differs in some essential but hidden
way from the outer layer, in kinds of fatty acids or something else,
mirroring the opposition between space and mass, between forces
FA and FG? There is
the above mentioned inversions in directions that nets of P-lipids
can undergo with P-groups inward or outwards. (When? Under which
conditions? If it is or gets explained, what could it eventually
tell us about this relation FA - FG
in macrocosm?
Perpendicular to direction of a lipid, along the
membrane,,,, L-waves seems expressed in motions:
There is the curious fact that in the fluid phase "a given
lipid will exchange locations with its neighbor millions of times
a second" and thus can wander along the membrane (Wikipedia:
Lipid_bilayer). Is this behavior really necessary! Biochemically?
It sooner seems as just an illustration of an L-wave → ← → ← → ← and the pole exchanges in d-degree
0/00 of Motions in the dimension
model.
In the model we have structure of a certain d-degree
plus debranched d-degree as external motion. Regarding the fatty
acids as substantiated structures of L-waves, the debranched d-degree
- in a step from higher Density ( 5 →>
4) to lower - could get this expression?
About proteins, they should illustrate the
relation between FE and FM,
the electric and magnetic forces or factors of FEM.
Besides their role as cytoskeleton, like a static EM-field, they
may be said to represent a communication system, distributed long
ways as are EM-waves.
We could ask if any of the many polarities between
amino acids or their codons in some way represents the opposition
M-fields versus E-fields with some functions related to circular
versus radial geometries? Surely an odd and here unanswered question.
It's easier to imagine the relation between purine
and pyrimidine bases of codons in terms of E- versus M-components
(G,A versus U, C or ?).
About amplitude versus frequency modulations:
Isolating the features of inward - outward directions in membranes
versus proteins, it may be asked if the polarity between amplitude
and frequency modulation in some way could be applied on these substances. In
atoms the absorbed incoming energy of photons get translated to
higher, "circular" amplitude of electron orbitals, while
frequency of emitted energy are decided by radial electron jumps
between orbitals. In nervous signals we have the similar polarity:
amplitude modulation of incoming signals through dendrites of the
nerve cell, note spread in its membrane, and frequency modulation
in outgoing responses through its axons towards other cells.
The genetic code is a "language" and
the 20 classical amino acids correspond in number roughly with number
of phonemes in different languages, phonemes which are different
combinations of frequencies.
Lipid membranes are difficult to study and hitherto
they don't seem to be attributed much of individualized properties
by scientists. However, the different membranes of nucleolus, the
nucleus, the endoplasmic reticulum, the Golgi apparatus and
cell membrane represent roughly different amplitudes. Could their
positions eventually give them some hidden but essential differences
that decide their roles for different processes and metabolism?
One example is perhaps that certain parts of ER attract ribosomes.
A more obvious one is the Golgi apparatus, where protein chains
are undergoing different modifications within the different apartments
outwards: why these steps in the modifications?
 Elementary
sketch of membrane amplitudes. Fig 23-147-2 Elementary
sketch of membrane amplitudes. Fig 23-147-2
9. A cell as substantiation of motions:
In a dimension chain of structures outwards, the debranched degrees
transformed to external motions come to form a dimension chain in
opposite direction inwards. This according to the elementary postulates
in the dimension model.
It awakes the thought that a cell would be best
understood as a substantiation of this inward directed chain of
motions, in this sense also an "inversion" of the atom
(cf. No. 4).
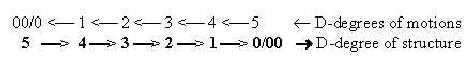 Fig
C-24 Fig
C-24
Related is the fact that most structure-building in the cell occurs
as steps inwards, from single units to linear additions to 2-3-dimensional
structures of the type 3←
2←1←
0.
Among the multitude of motions there are the high
mobility of cell membranes, invaginations and evaginations as 3-dimensional
motions (~ convex/concave as one
polarity of d-degree 2) and contractions/relaxations
of protein threads as a kind of 1-dimensional vibration.
Nucleus and other organelles of a cell can also
move around in the cell (cf. "translational motion in 3 directions?)
and even rotate. The cytoplasm itself is continuously streaming
in different directions (Fb p. 10).
The whole cell as a prison for trapped motions?
The metabolism of cells with nourishment from its environment
corresponds to the presumed "breathing" of atoms, i.e.
how atoms uphold their existence and structure through "breathing"
of what is antimatter on their level - the surrounding "empty"
space. Structure of matter breaks down when gravitation of Mass
chokes the access to Space. Cf. the interpretation of light
waves too.
10. About number 5 - as dimensions in the model:
- One, most elementary example is valence 5 of the P-atom: the
HxPOx-groups having organizing and energy-storing functions in the
cell.
(Could inorganic phosphorus eventually have served
like a grain for growth of snow crystals, when a cell appeared?)
- A second example is ribose, the 5C-molecule of carbohydrate,
essential in .nucleotides. As mentioned above the Pentose-phosphate
cycle transforms 3 ribose C5 to 5 C3-molecules. There we have the
number 15, sum of the d-degrees in the dimension model Cf. HCOH
= 30 A, sum of poles in the model.
- Another 5C-molecule is isoprene, three of which join to
one C15 and these may join head to head, i.e. in opposite directions
to form squalene of C30 on the way to steroids; 30 the sum of outer
complementary a- and b-poles in our model.
Mentioned above too is the old, single-cell Archae
bacteria, which uses isoprenes to create fatty acids (Wikipedia).
As two C15 molecules can meet head to head, so can these isoprenes
of Archae join their tails in a similar way.
- On the level of organelles it has been found that ribosomes,
where protein synthesis occur,
appear usually in groups of about 5 in bacteria (Bc p. 79).
Ribosomes are also very old organelles, compounds of RNA and proteins
(cf. figure 1!).
- The Golgi apparatus in eukaryotic cells is described as most
often consisting of 5 - 6 christae (Aph), membrane
enclosed apartments or factory sections.
- Another kind of example could be the number of enzymes, 5,
involved at copying of DNA to mRNA, a process demonstrated by Roger
Kornberg and his team at Stanford about 2007.
- Finally we could remind of the 4 - 5 cyclic processes
that scientists (e.g. Marquand) have considered as necessary
conditions for life.
About cancer, some very abstract thoughts.
Development of cancer has been said to occur in 4-5 steps.
 Fig
C-25-154-1 Fig
C-25-154-1
0-pole is defined as primary, integrating binding force in the
dimension model, 00-pole primary polarizing force. The 0-pole is
also the one of unity, the 00-pole the one of multiples.
Experiments with "exogastrulation" in development of
embryos
showed that the cells of the animal pole (~ 00-pole) when tied off
from the vegetative pole only developed to an amorphous, hollow
mass, while the cells from the vegetative pole (~ 0-pole) differentiated
towards different organs, although not complete.
Cancer as amorphous, undifferentiated cell divisions
could thus correspond to the result of an isolated 00-pole, its
uninhibited, unregulated divisions when the 0-pole as integrating
binding force has been weakened.
The observation that cancer cells often first
appear in rings (Aage Möller 1974, lecture) seems to
support the analogy with exogastrulation. It's also said that pregnancy
can have a suppressing effect on cancer - perhaps because an embryo
represent a vigorous center.
A 0-pole, a center, gives outward direction as integrating force,
which could correspond to field lines from a cell outwards towards
other cells. A 00-pole, an anticenter, gives inward direction as
a polarizing and isolating force.
Mutagen, carcinogenic chemical substances contain
for instance often CHx-groups which give inward direction (aromatic,
ring-shaped carbohydrates too, in molecular structure inward directed).
Cf. for instance the T-base = U-base plus a CH2-group
that turns a strand of nucleotides inwards to DNA. Characteristic
for cell division in opposition to interphase is duplication of
DNA with T-base instead of U as in mRNA.
A cancer cell leaves the tissue level of organization
and 'casting', the distribution of roles, and returns to the cell
level. In the big chain of levels this is also a direction inwards,
towards underlying level compared with the cell as part in a bigger
whole.
In this respect cancer would be about the same
as the relation between unicellular and multicellular organisms.
(Which proteins or genes separate these classes?) Cancer can be
encapsulated like "resting spores" - and spread as "swarm
spores", leading to metastases in the body.
It's said that similar, repeated stimuli or irritations from outside
- of all kinds - can initiate cancer, one-sided pressure and even
distilled water repeatedly brushed on laboratory animals. This could
be translated to a suppression of "the whole" and with
it the integrating center in the organisms? The organism's response
to the stimuli may lead to exhaustion that in reality depends on
"all the rest" that doesn't get activated, the whole that
are dismissed in favor of the part.
11. Prokaryotic - Eukaryotic single-cells:
Here about theories of how eukaryotic cells emerged. Other aspects
in additional file
(No.. b) to Aviation,
Prokaryotic unicellular organisms (PKc) as blue-green algae and
bacteria lack nuclei with membranes and mitochondria. Eukaryotic
cells (EKc) have both these membranes and several other organelles
in cytoplasm.
PKc have similarities with the embryological
blastula stage of multicellular organisms, EKc with the
gastrulation stage with several inner tied off parts of membrane
à la "vesicles" to creation of specialized tissues
and separate centers.
Compare too the evolution
from 1- to 2- to 3-layer organisms.
It's a common view on embryological development that it reflects
earlier stages of evolution. We could presume that the same principles
for 'evolvement/involvement' are
prevalent on underlying levels as on superposed ones.
Thus, it becomes a possible hypothesis that development
from algae and bacteria to EKc occurred in a corresponding way,
through invagination (of material) of their cell membranes.
But why should it take billions of years?
Another hypothesis among scientists is that mitochondria and chloroplasts
are bacteria and algae that have immigrated into or become incorporated
by bigger, heterotrophic cells - and then lost their capacity for
independent existence. (According to some theory the nucleus itself
should have been an outsider that had immigrated into another PKc
before chloroplasts.).
Similarities between ribosomes in EKc and PKc
have been one reason for this hypothesis. However, it doesn't explain
emergence of the nuclear membrane in EKc. It's also pointed out
that several components of mitochondria "curiously enough"
are synthesized in cytoplasm of EKc outside mitochondria.
Such things seems sooner to support the other
hypothesis among scientists about invaginations of own membrane
of the gastrulation type. Similarities in ribosomes could be regarded
from the aspect that organelles of PKc and EKc make up analogous
dimension steps on different levels. And cilia have also a little
DNA but are not attributed or suspected to have had any earlier
independent existence.
There are other facts that seem to favor the theory about invaginating
cell membrane. There can be found such invaginations to "mesomeres"
already in PKc and certain structures analogous with an endoplasmic
reticulum (Bc p. 290). The theory needs of course another
explanation of DNA in mitochondria and chloroplasts, but already
in PKc smaller, ring formed DNA-strands (plasmids) exist outside
the bigger DNA-thread (Fc p. 168 f.).
The relation between EKc and PKc has features of the type center
— anticenter, also the one between higher and lower d-degrees
but this relation is ambiguous. Certain features point to PKc as
of higher d-degree in our interpretations: the use of both L- and
D-forms of amino acids in their membrane proteins, the less polarized
(not sexual) cell divisions and the less amount of inner motions
for instance (see here).
In other respects PKc have the features of anticenter. They remain
a multitude of single-cells, while EKc leads further to multicellular
organisms, to a new level. (Cf. the small satellite cells
in he peripheral nervous system surrounding the bodies of big nerve
cells.).
This ambiguity could follow from a dimension chain,
where debranched degrees in first higher steps meet "the other
way around" from anticenter, the 00-pole, inwards.
A general postulate or aspect on the dimension
model for evolution is also the stepwise building in of the 00-pole,
the anticenter.
Perhaps the two different hypotheses above can be seen as only
two aspects on the same development, only two opposite directions
of reading a dimension chain, illustrated in the figure below.
1) Prokaryotic cells as created from debranched
degrees in higher steps, meeting the other way around in synthesizing
direction inwards 3 ←
2 ← 1←0/00, immigrating into cells
from higher d-degrees: a reading of the chain inwards.
2) Pre-eukaryotic cells from higher d-degree steps
5 →> 4 →>
3 meet at their outer surface (d-degree 2 ) structures from steps
2 ← 1←0/00, which get enclosed
through invaginations. This is a more or less equivalent description
than that in point 1, reminding of the gastrulation of multicellular
embryos from blastula stage.
An ambiguity appearing in step 3 - 2.
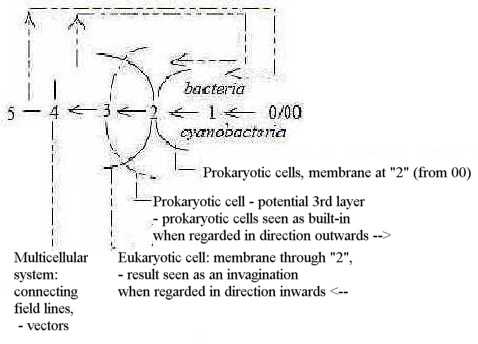
Fig C-26-162
The similar ambiguous relations appear among multicellular
organisms when it concerns 1-, 2- and 3-layer animals.
The two theories become connected with the two ways to look at
the dimension chain, "horizontally" as a "straight
chain" in synthesizing direction inwards - or "vertically"
as in the loop model, outward - inward directions meeting in step
3-2: d-degree 5 polarized a) 0-00, b) 4-1, c) 3-2.
Should this view imply that EKc perhaps existed much earlier in
time than believed, that both kinds EKc and PKc eventually emerged
as c-ac-poles simultaneously? In a relation one to many? Archae
bacteria, now regarded as an own phyla, show certain features
similar to both PKc and EKc.
One question is why only one (or some?) PKc transformed to EKc.
A special change in outer environment? A mutation? Something that
got wrong at cell division …? A kind of "neoteny"
in the evolution of chromosomes? - Or just an underlying principle,
implying steps towards more complicated structures as in a dimension
chain, in similarity with how classes of substances develop from
strings towards higher d-degrees in shape: 3 ←
2←1 ←0?
Invaginating layers and cells as trapped light?
A related speculation concerns the layers of different substances
that surround cell membranes of many organisms: outer layers of
proteins, glycolipids, mucopolysaccharides and such combinations
of the more elementary classes of molecules.
Could perhaps such layers have been a first phase
during evolution? (Compare the hexagonal pattern in first figure
above) Layers that invaginated and closed and made outer kinds of
molecules inner ones? A pole exchange center - anticenter in our
model. This in the same way - and by the similar forces - as the
blastula transforms to gastrula in multicellular organisms?
In the dimension model, as said above, one general aspect is the
stepwise "building-in" of the 00-pole.
And what about strings of nucleotides? It's a bit curious that
DNA as chromatids on photos seem mostly attached to the inside of
nuclear membrane. It's said that DNA-thread in bacteria is attached
to their cell membrane. Couldn't DNA also primarily have been formed
on the outside of a membrane layer, exposed to light, and then have
invaginated? Chromosomes as molecules for refraction of light could
on an early stage of evolution easily have been affected by chemical
environment at a surface? Chromosomes as "substantiated light",
"lumosomes"? (Autotrophic bacteria that used other energy
sources should in that case have been a later adaptation?)
The light-capturing chloroplasts with DNA are
for instance located along the surface layer of the cell wall in
Spirogyra algae, and development of chloroplasts is dependent on
light exposition. The respiratory chain in mitochondria with DNA
is attached to its cell membrane.
Steroids get bound to lipid membranes but binds
also to DNA.
With the mentioned facts it seems possible to imagine that DNA
(or rather RNA) once were located and constructed on the outside
of a lipid membrane and then through invaginations became the inside
- immigrated. Cf. on the multicellular level invagination of the
neural tube: outside becomes inside, environment built-in. Cells
as trapped light?
Cf. Evolution,
the turn inside-out of blastula in embryology of some sponges and
similar processes in colonies of flagellates.
12. Some final words:
What could have been the real root of a first cell? We may speculate
about an original conglomerate, a multidimensional network, something
like the principle in the hexagonal pattern in figure 1 above, where
different stereometric configurations and points of refraction in
different angles and readings of patterns - or restratifications
- have given the different classes of substances and combinations
of them. And that meshes in the network as matrices or the like
could have been the origin of the coupling between proteins and
codon bases, bases where parts of the rings simultaneously make
up atoms in amino acids. Structure relations in the network then
through polarizations and substantiations developed to processes
of syntheses for different substances.
Yet, what could have served as a very first "center"
and first "anticenter", poles within which a cell developed,
in a sense that agrees with the dimension model?
An eventual complex network or series of layers
is hard to see as such poles out of some 5-dimensional pre-unit.
Nor does DNA resemble such a first center.
- Just energy in some form, serving as center? Corresponding
to how amino acids and bases etceteras on an underlying level are
shaped from elementary molecules in Miller-Urey type of experiments?
- Or one single hydrogen atom H, the inversion of which appears
as first anticenter? As guide for a pure mathematical, organizing
principle underlying the development? (Cf. about quotient e/p
in files here
and here
about the genetic code. About inversion also file
1/7; 7 "not developed"
d-degrees according to the string theory.
- Or a P-group, inorganic phosphorus (H3PO4),
an atom as P with valence 5 as core for condensation, functioning
as the speck of dust for an ice crystal around which it gets organized?
- Or just organic molecules trapped in a metal environment as first
anticenter? The polarity of metals - non-metals, defined by the
"octet rule", enough as definition of first center-anticenter
poles?
…
A center defines directions - outwards /
inwards. In chemical terms for instance hydrophilic - hydrophobic
directions. In next step in the model a spherical structure
versus a radial one gets defined.
Within the cell as a whole, a unit, interpreted
as a whole dimension chain, each step can according to primary postulates
develop to whole secondary chains, the steps in these to tertiary
chains etceteras.
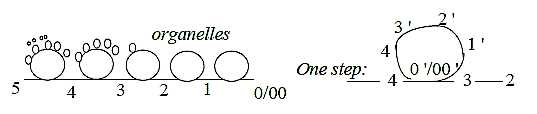
Fig C-27-143-2
It gives an exponentially increasing number of directions and structures
within the cell and increasing number of motional moments as processes.
The hypothetical conglomerate of both lipid layers
and nucleotides and proteins could then be imagined to polarize,
creating increasing distances within these poles, with development
of circular processes from the different steps. Something like the
embryological gastrulation of a eukaryotic, multicellular organism.
*
A note:
It's hard to believe that all cells on the Earth derive from just
one single cell, even if it's unknown how a cell is born in other
ways than through duplication. Why then look for life on other distant
planets! It's illogical.
Either there exist a principle of Nature of only
one single cell as one center of Universe (which happened to be
here). Or life appeared under right circumstances on a lot of places
- on the Earth and other planets.
To an extra link: Levels
from Micro- to Macrocosm
|