|
1. Relation between organs for equilibrium and hearing:
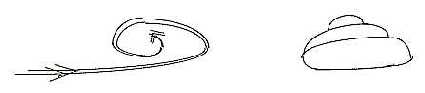
Fig
H-1-117-1
We have assumed that gravitational waves, if they exist, are of
the longitudinal, linear type: → ←
→ ← → ←
So are sound waves, conveyed through variations in pressure on the
tympanic membrane in cochlea. Pressure is a quantity F/m2,
the force F here an inward directed one as is gravitation. Thus,
it's rather natural that the sense of hearing is developed in close
relation to the sense of equilibrium, even though it sometimes has
been called a 'mystery'. (Cf. 'pressure' as increasing 'Density',
proposed as only term for first physical quantity defined in step
5 - 4 in the model here, before gravitation gets defined in next
step.)
The organ of equilibrium
concerns own position and movements of the individual, in this sense
referring to outward activity from the organism as center, a 0-pole.
Hearing is primarily an organ for input, impressions from outside
the environment, from the 00-pole. Hence, the two organs may be
interpreted as representing a polarization in directions outwards
→> ←
inwards, which may be one factor behind the differences in developed
structures.
Hearing is also "time displaced", later developed during
evolution. A small canal from saccula in the organ of equilibrium
develops to a tube which grows during evolution, becomes a bent
tube in reptiles and then the convoluted spiral in cochlea of mammals
(figure above).
In the dimension model the outward direction for dimension degree
(d-degree) of structure: 5 →>
4 →> 3 →>
2 →> 1 →>
0/00 gives the opposite chain from 0/00
stepwise inwards to 5 in d-degrees of motions. The
development of the organ for hearing from first a "linear"
tube to rotating spiral could be apprehended as a substantiation
of the pattern of motions of increasing d-degrees. (In opposition
to the structures in organ of equilibrium, from sacs as volumes
to half circle bows with functional steps towards increased d-degree
of motional reaction from linear to rotational movements.)

Fig H-2-117-2
2. Number 3 in structure appears in cochlea too:
- Spiral turns - like shell of a mollusk - are nearly 3. In cross-section
it gives number 5 in-out as 1-2-3-2-1.
- The auditory ossicles in middle ear are 3 in mammals (Reptiles
have only 1 but already among ray-finned fishes one finds 3 small
bones on each side of the 4 first vertebra that convey pressure
changes from swim bladder to the labyrinth (Kz).
- The inner of cochlea tube becomes divided in 3 canals.
3. Cochlea and organ of equilibrium as one dimension chain:
In the spiraled tube of cochlea the anticentric canals with perilymph
are those in which the pressure waves from outside are transmitted
in - out. The middle, central canal with basilar membrane contains
enclosed endolymph, the same as in the sacs of equilibrium organ.
Cochlea and organ of equilibrium are joined through
the mentioned very small canal for endolymph from saccula to the
middle canal of cochlea.
If we identify parts of the equilibrium organ
as expressions of d-degree steps, saccula with step 5 →>
4, then the small connecting canal could be identified with the
debranched degree in that step: hearing as derivative of static
pressure formed by that debranched degree in opposite direction?
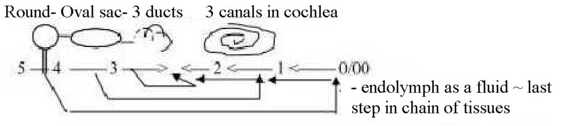
Fig
H-3
Cochlea as from debranched d-degrees.
Figure above broken to positioning of right part straight above
the left:
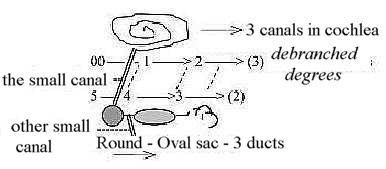 Fig
H-4 Fig
H-4
Two other small canals connect endolymph and perilymph respectively
to the brain:
- One departing from between saccula and utricle (as in step
4 →> 3, marked in the figure
above) for endolymph, going to the dura,a layer near
the space for CSF around it. (Cf. CSF departing as a side branch
from 4th ventricle to circulation around the brain.)
- One, containing perilymph, departing from cochlea (?, divergent
information in sources), going parallel with the preceding one to
the subarachnoid space between membranes of the brain, in closer
contact with the CSF liquid.
The three canals seem to illustrate ramification
(polarization) of degrees and join the two organs as to one dimension
chain.
If the ducts of equilibrium organ only contains perilymph (according
to Nf p. 396) or, which seems more probable,
in similarity with sacs and cochlea includes endolymph too as later
illustrations show, is left here as an open question.
It seems possible also to describe the opposition between
the two organs in terms of centric - anticentric lymph:
- It's motions of the centric endolymph that directly activate the
hair cells in organ of equilibrium.
- It's pressure waves in the anticentric perilymph that Via a membrane
effect the endolymph and hair cells in cochlea.
4. Oval - round window and upper - lower canals:
Pressure waves from tympanic membrane are conducted to the oval
window into perilymph, upper canal. Through a small hole in apex
of the cochlea the pressure can turn to outward direction in perilymph
in lower canal and to the round window.
Here is once again the polarity between round
and oval forms, as a step of polarization one to two focal points,
connected with the polarity outward - inward direction (and ultimately
with 0-00-poles):
- ac - oval window →>
inward direction - upper canal.
- c - round window←outward
direction - lower canal.
"Upper" versus "lower" as between distal versus
ventral sides in the whole nervous system. This latter polarity
is defined through the features of the middle canal:
- basilar membrane with hair cells on the wall to lower canal,
- the flap of a membrane (tectorial membrane) that lies over the
hairs and affect these from the wall to upper canal.
5. Cochlea as illustration of forces:
a. It's the difference between the pressure forces inwards
on the oval window and outward on the round window that decides
the effect on the receptor cells in the middle canal (AM),
hence a kind of derivative.
Suppose that we associate the force in pressure
(F/m2) inwards on the oval
window as derived from gravitation (FG)
and the opposite direction of pressure outwards on the round window
as derived from the outward acceleration force
(FA). The change of directions inwards
- outwards in the small hole at apex of cochlea have the character
of a "pole exchange", presumed occurring in last d-degree
0/00, equivalent with 5' in the
dimension mode. (Cf. what happens in the "bottom" of black
holes!)
From apex an d-degree 0/00
of motions the growing spiral of cochlea illustrates the substantiated
4-dimensional motion assumed in the model of a linear 1-dimensional
structure.
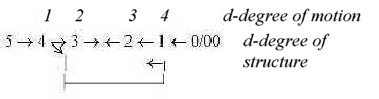 Fig
H-5 Fig
H-5
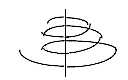
b. Association to magnetic fields - and d-degree of motions:
The whole cochlea along the length axis gives in cross-section the
picture of anticentric motion of canals and pressure waves around
the central axis with its nerve fibers from the hair cells.
 Fig
H-6-118-1 Fig
H-6-118-1
It reminds also of the magnetic field around an electric cable.
In files about forces
and electromagnetic
waves the magnetic factor is suggested as "the son
of" gravitation in the following d-degree step 3 - 2. (FG
as one of the two forces in d-degree 4 and step 4→> 3.)
Then, a magnetic field around the electric nerve
fibers could be partly responsible for the structure of cochlea?
However, the graded radii of circles demonstrate
simultaneously what we in file about motions
have suggested as a 4-dimensional motion in d-degree 1
as "pumping".
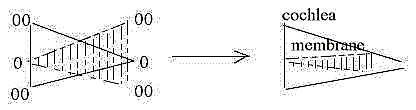
6. Two gradients:
The form of the bony cochlea, from the outer broad base towards
its inner apex corresponds to inward direction from anticenter to
center, 00 →> 0. The basilar
membrane with hair cells in its middle canal has the opposite structure:
most narrow at the broad entrance of cochlea, widening toward its
inner, narrow apex as direction 0 →00.
It gives the principle structure of two opposite gradients in each
other:
 Fig
H-7-117-3 Fig
H-7-117-3
Breadth of the basilar membrane furthest out in relation to furthest
in is 1/5 (Nf) - which perhaps
could be seen as expression for relations in a dimension chain?
The construction feels odd* with regard to space
but reveals and underlines the clear, complementary polarity of
type center —
anticenter, receptor structure versus the surrounding bony cochlea..
* (Is explained by the smaller size of central
axis nearer apex.)
The figure of gradients could be compared with directions in
the production of lymph, fluids in opposition to
structures a phase relation of the type 00 to 0:
- The endolymph is produced and secreted by specialized cells
in the middle canal of cochlea (Aph p. 578): hence in direction
inwards with the dimensional interpretation above, from organ
of hearing to that of equilibrium.
- Perilymph has the similar content as CSF and it would be logical
to presume that it derives from the other end, outwards, perhaps
from CSF in the brain? (No information available.)
It should imply opposite directions of currents
in accordance with the c-ac polarity but as motions of fluids
reversed in directions in relation to structures of the organs.
A similar reversal appears in relative charge
compared with the inner of a cell and its extracellular environment:
Central endolymph is positive (+80 mV) in relation to the anticentric
perilymph. It doesn't depend on the ion balance; endolymph as
central contains much K+ as the inner
of cells, perilymph much Na+ as the fluid
outside cells, but here it's the perilymph that contains most
proteins, responsible for most negative charge within cells.
Both these reversals could be regarded as results
of the opposite directions in the structure according to the suggested
interpretation - and most likely as phenomena on different levels.
From the picture of two gradients it follows that basilar membrane
with divergence inwards has its origin at anticenter of the body.
Generally, receptor cells represent the inward direction in the
main polarization of the nervous system in the sensory - motor pathways,
thus could be said to have their starting point as 0-pole furthest
out at the surface. (They derive obviously also like
the organ of equilibrium from the lateral line system of fishes.)
In this sense the anticenter becomes built-in into center, the central,
endolymph canal in cochlea.
The bony cochlea becomes the inverse, produced
from inside out from temporal skull bone. Diverging outwards it
encloses the basilarmembrane and inner canals.
Such a feature of design, where tissue material
from inside becomes anticentric to invaginating material from outside,
recurs in several cases in embryology
(No. 10 c).
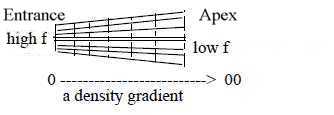
The basilar membrane has its highest density as stiffness at its
"0-pole", nearest the entrance, making it a gradient of
density. Cf. "density" regarded as first physical property
defined in d-degree step 5 →>
4 in our model.
 Fig
H-8-121-2 Fig
H-8-121-2
Highest frequencies are registered nearest the entrance, the "0-pole"
of membrane as gradient.. It's partly depending on the higher density
of membrane here. Cf. EM-waves where higher, more energetic frequencies
originate from center of an atom. Long wavelengths, lower frequencies
reach their maxima further in towards apex where membrane are broader.
In musical terms we get the deepest "fundamental tones"
at apex of the cochlea, the center of other gradient, its "overtones"
further out in cochlea; in this apprehension in opposite direction
from deeper levels to superposed. What's fundamental and depth must
obviously here be seen as decided by the waves in perilymph, not
the membrane in itself.
In agreement with this latter aspect, signals
from apex (lower tones) are registered by centers in the brain at
the ventral side, signals of higher frequencies from the
entrance at distal side, sides corresponding to 0- versus
00-poles in the embryo (Nf p. 409).
7. Arrangement of hair cells:
The arrangement of hair cells on the basilar membrane seems to reflect
the construction of the whole cochlea: 3 outer rows of hair cells,
with cilia in V- or W-form turned inwards the central axis, as mirroring
the 3 spiraled canals, and one inner, linear row of cells with linearly
arranged cilia along the central axis with ganglions and nerve fibers.
In cross-section of the cochlea it appears as one version of the
radial / circular poles 3b-3a in
the dimension model.
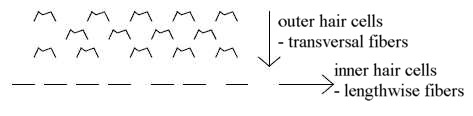
Fig H-9-119-1
Cilia, the "hairs", of the outer cells are also arranged
in 3 rows on each cell, in 2 rows on inner row of cells. Number
of cilia on each of the outer 3 rows is about 100, on inner row
about the half, a relation ~ 2/1
(Nf p. 398).
Outer hair cells are more sensitive to motions inwards the central
axis, the inner ones more sensitive to lengthwise motions along
the axis (Nf p. 399): another expression for the same geometry.
There are also nerve fibers of two kinds, transversal ones from
outer to inner cells and to the central axis, and lengthwise spiraling
ones.
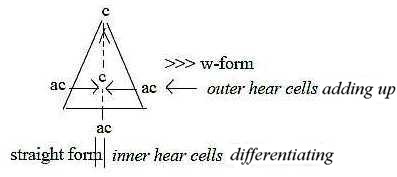
The polarity center - anticenter is expressed in many ways.
- It seems as if the outer hair cells "activate" the inner
ones (AM), if so just as well as it is the pressure waves
in anticentric canals that activate all hair cells. It should be
logical with the sensory nervous system as inward direction and
anticenter pole as the polarizing force in our model.
- A further example is the polarity converging - diverging signals
in the coupling of nerve fibers: signals from about 10 outer hair
cells converge to one nerve fiber (convergence from anticenter),
and the signal from each inner hair cell is spread to about 20 afferent
nerve fibers (Nf p. 402), i.e. diverge. (Divergence from
a center pole.)
These relations should imply that outer rows of
cells summarize impulses over a broader part of the basilar membrane
and that it chiefly is the inner ("linear") row of cells
that discriminate between frequencies in sounds.
It looks like the polarity in every nerve cell
is transformed to this whole multicellular system: amplitude modulation
(summarizing) of incoming signals and frequency modulation of outgoing
signals: principally perpendicular entities.
 Fig
H-10-119-2 Fig
H-10-119-2
8. Wave forms:
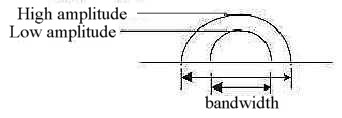
Frequency and amplitude as complementary energy forms become translated
in different ways in the pressure waves and basilar membrane:
Higher amplitudes increase the bandwidth (~ lengthwise)
of frequencies, however most for high frequencies (Nf p. 408).
Low amplitudes give more narrow maxima. It could illustrate the
principally perpendicular relation between these forms of energy
as between circular and radial poles out of d-degree 3, originating
from anticenter and center respectively in our model.
An illustration of the principle:
 Fig
H-11-121-4 Fig
H-11-121-4
The increase in bandwidth at high amplitude of sounds and the inverse
at low sounds could be compared with the relation between high and
low temperature, another form of energy that seems analogous:
High temperature (to compare with high amplitudes
of sound) corresponds to great spread of particle velocities, low
temperature to more equal velocities of the particles. Principally
it gives heat and cold as properties at straight angle to one another:

Fig
H-12-122-1:
As mentioned above, density (~ stiffness) as a factor behind the
frequency distribution on the basilar membrane is in the dimension
model proposed as first "physical quality" defined in
d-degree step 5 - 4. Temperature as motion of quanta is as "physical
quantity" defined in last step 1 - 0/00.
A correspondence seems natural with the loop version of the model
in mind.
About frequencies (f), there is the other polarity between
high and low f:
- Long sound waves (low f) have long rise times, i.e. reach
their maxima further in, nearer apex, but have steep, short fall
times.
- For short waves (high f) it is the reverse: short rise
times, longer fall times. (High f at the entrance = "0-pole"
of basilar membrane.)
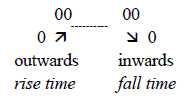 Fig
H-13-122-2 Fig
H-13-122-2
Rise and fall times correspond in this way with the main direction
of basilar membrane in the illustration of gradients above.
9. Inhibition.
Inhibition of the lateral type between receptor cells isn't found
in the cochlea. There are instead efferent nerve pathways from higher
nuclear centers in the brain whose axons have synapses with the
hair cells. (It seems to imply that hearing is an active, discriminating
process!) This "antiparallel" inhibition from higher centers
could in terms of the dimension model show hearing as a sense of
higher d-degree than for instance sight
with lateral inhibiting cell layers - or just on hearing
as a later sense in the history of evolution, geometrically less
developed?
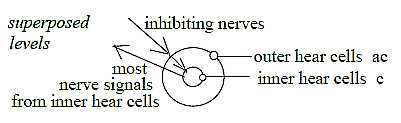
Inhibiting nerve fibers from higher centers go
to the outer rows of hair cells (TA p. 88), whose activation
of inner row of cells consequently should be hampered. If so, an
example of indirect inhibition in two steps, inwards in a level
chain:
Fig
H-14-120-2
In the sensory system as inward directed, the outward directed activity
from inside, (fundamentally associated with the motor system), thus
becomes inhibiting. In its function also serving contrast:
Activation through these efferent pathways on
tones just above or under a certain tone can have inhibitory effect
on the frequency of this tone (AM-Hf), in this sense an indirect
"lateral" inhibition between receptors.
Certain cells in cochlear nuclei are inhibited
by tones with frequencies on both sides of its own frequency, other
cells only by frequencies on one side, above or under its own (Nf
p. 410), a differentiation corresponding to further polarizations
and increasing "one-way-direction". (Cf. similar polarizations
among on-off-cells of ganglia in retina.)
(Hearing impairments often occur ½ - 1 octave above
the frequency of the injurious sound but can also spread to lower
frequency areas (AM-Mb). Thus, it becomes a natural question
if such injuries depend on too strong (killing) inhibiting activity
from inner, higher centers (?). 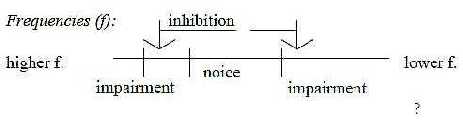 ) )
Fig H-15-121-1)
Another observation is that signal answers from hair cells depend
on velocity of changes in stimulating tone frequencies. Fast changes
give higher, narrower spikes, more distinct discrimination of frequencies
(Nf p. 412) in cells of higher centers (nucleus cochlearis).
It shows on their property as derivatives, developed in many senses,
one feature that reveals d-degree steps - as in mathematics. (Velocity
tentatively presumed in the model here as the physical entity for
the very d-degree steps, distance/time.)
10. Number of hair cells and nerve fibers:
Data vary but one reference says hair cells in a human ear are ca.
15000, nerve fibers from cochlea 25000 - 30.000 (Nf p. 398, 400),
thus up to twice the number of cells. (Ca. 12000 in outer rows,
3000 in inner row, a relation 4 to 1 - or 4 to 3 in individual
rows of cells.) The number of cells happen to be ~ 103
times the sum of a dimension chain, number of nerve fibers sum of
the poles in this chain:
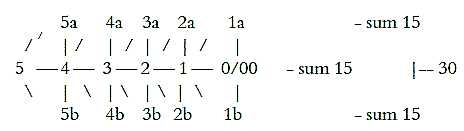
Fig
H-16-120-1
|