|
1. Some general aspects:
Its hard to see how only "random mutations" and
"natural selection" could explain an evolution towards more complex
organisms. All kinds of variations, yes. But hardly the increasing
"capacities" of creatures from the enormously well adapted bacteria.
It has been said that even the simplest unicellular
forms of life that have survived to our days have nearly all types
of enzymes as higher animals (JB), the same number and kinds
on different levels in the evolutionary chain. And all fundamental
functions exist already in the unicellular organisms.
If this still is valid
after an immense amount of new knowledge during last decades, it
has implications for the Darwinian view on evolution.
When it concerns multicellular organisms all phyla
within the animal kingdom seem to have been differentiated already
in Precambrian times (Ez). "In no case any transitional forms
of today or as fossils are found."*
*If now this statement isnt a kind of circular
definition: the definition of phyla builds probably (?) on the
lack of transitional forms. The classifications of phyla are
also much debated.
For instance: fossil finds cannot confirm an evolution
from 1- to 2- to 3-layer animals. On the other hand an evolution
in the dimension of time seems proved within a group as chordates.
If we assume (irrespective of divisions into phyla)
that all basic forms with different degrees of complexity appeared
more or less "simultaneously" from the eukaryotic cell, we have
to imagine that they got differentiated out of some underlying,
basic differentiating principles, perhaps as through a series of
more or less momentary quantum jumps; the following differentiations
within the basic forms along another coordinate axis of time. This
as analogous to the first seconds of Universe after big Bang with
following development through billions of years.
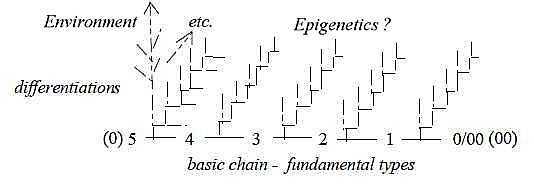
Fig
Ev-1-177
Environment changes slowly. If crediting Darwinian
"trial and error" mutations 50 % of the truth, we have still 50
% to account for and ascribe to basic structural principles.
Then, the longtime evolution
should be connected with a hierarchy of genes of the same kind,
with primary rows of genes differentiated into secondary, more specialized
ones; the primary genes perhaps getting activated at certain threshold
points of major changes in evolution?
The different coordinate
axes may also be seen as appearing in the relations egg →
individual versus individual → environment:
egg the 0-pole, environment the 00-pole. Its
a bond of dependence that implies mutual changes between the "poles".
2. Biologists primary bases for classification
of organisms:
Sub-cellular level:
a. Autotrophic — heterotrophic
organisms
(= not depending
depending on organic nourishment as plants versus animals)
b. Prokaryotic — Eukaryotic
organisms
(cells without nuclear
membrane as algae and bacteria cells with nuclear membrane)
↓
Eukaryotic organisms, animals:
- Unicellular multicellular organisms
- 1-, 2-, 3-layer organisms
- Coelom (body cavity) types: degrees in development
of a true abdominal cavity
(Acoela, Pseudocoela, Schizocoela, Enterocoela)
- Protostomia Deuterostomia: oppositions in
directions regarding formation of organs.
(Deuterostomia is the branch leading to
vertebrates, mammals and Homo Sapiens.)
About classifications on the sub-cellular level, a) and
b) above, see additional
file.
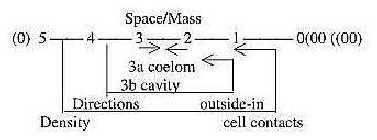
In the last 4 points of classifications
its easy to recognize the geometrical steps in dimension degrees
(d-degrees) of the dimension chain, our background
model. A rough, first description of the evolution in synthesizing
direction inwards in the chain:
- from single cells as points to cell-contacts of multicellular
organisms, a step 1 ← 0,
- to number of tissue layers as surfaces, ←
2 ← 1
- to separation of inner abdominal room in mass and space, step
3 ← 2,
- to the differentiations in directions inwards/outwards,
front-back etc. in step 4 ← 3.
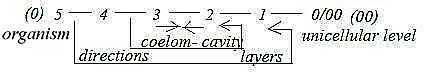 Fig
Ev-2-179-3 Fig
Ev-2-179-3
In the dimension chain in physical and geometrical
terms:

Fig Ev-3-179x
a-poles with features
from 00-pole, b-poles with features from 0-pole:
0-00 also a relation between higher and next lower d-degree.
The level
chain in a developed chordate corresponds rather naturally
to these basic classifications:

Fig Ev-4-179-1

Fig
Ev-5-179-2
The first three steps above of evolutionary classifications corresponds
to the embryonic development of a 3-layer chordate:

Fig Ev-6-179-4
[A couple of following differentiations
within Deuterostomia:
- Invertebrates Vertebrates.
- Vertebrates: egg-laying — mammals.
- Mammals: without amnion —
with amnion.
Subgroups within group Vertebrates
Tunicates → Lancelets
→ Cyclostomes →
(e.g. lampreys) → armored sharks
(Placoderms) → Cartilaginous fishes
→ Lungfishes →
Ray-finned fishes and bony fishes →
Coelacanths ("Old Four Legs") → Amphibians
→ Reptiles →
Birds // Mammals → Homo Sapiens.
Biologists have divided living organisms in 5 kingdoms (Fc p.
95), a number 5 that hasnt much to do with the 5-dimensional
chain tested here:Unicellular prokaryotic unicellular eukaryotic
plants fungi (mushrooms) animals. ]
3. Unicellular to multicellular organisms, step 1 ←
0/00:
The evolution from first prokaryotes to eukaryotes and then to
multicellular organisms took billions of years. Changes of the environment
and chemistry of the atmosphere of Earth are taken as main factors.
However, there seems to be a lack in the explanations
between this very slow, continuing change of the environment and
produced, outer conditions on one hand and the clear, "sudden"
steps on the level of the cells themselves on the other.
It could indicate the existence of certain thresholds
in values of parameters (light, oxygen etc. ), which function as
borders for marked, new constructions.
As said before there are many intermediate forms with cell contacts
(regarded as d-degree 1 in the level
chain) forming colonies of individual cells, also among
prokaryotes. It's however only eukaryotes that develop to multicellular
organisms, obviously representing a new degree of integration.
Some aspects on this transition could hypothetically be
- inversions of some kind?
- new center defined at meeting between individual units of equal
potential?
- a deeper center as start of cell division?
- just a reversed relation in relative amount of e.g. DNA and proteins
versus lipids?
About inversions:
- Multicellular organism represent unity among cells - as guided
by the integrating 0-pole in our model. Unicellular organisms express
separation between cells - as out of the polarizing 00-pole. The
multicellular represent unity, the unicellular the multitude - in
spite of amount in their names.
The relation unicellular egg - developed organism
in the multicellular organism appear as a kind of reversion - or
the previous opposition built in: it's the egg that has the high
potential for differentiation, while the developed organism represent
already differentiated cells, corresponding to start and end of
a dimension chain as steps of polarizations.
In terms of directions outwards /
inwards it could be pointed to the circumstance that unicellular
organisms have a tendency to seek themselves inwards multicellular
organisms, to live inside these - as for instance algae in fungi
and bacteria in animal stomachs - like viruses more explicitly.
(A parallel to the theory
(No. 11) of mitochondria and chloroplasts as immigrants
in eukaryotic cells.)
Certain 1-layer species as calcareous sponges have
a "curious" turn of the blastula inside-out at their embryonic
development. Similar processes occur in colonies of flagellates
as Volvox (Ez p. 57), hence among individuals referred
to step 1 ← 0/00 in our interpretation
here, single cells to cell contacts.
Such "inversions" could be the expression for
the "pole exchange" in last step 1 →
0/00 of our model (where motions outwards
define a new anticenter, motions inwards a new center. (This last
step 1 0/00 is represented in each higher
d-degree step too.)
The invagination of vegetative
pole in embryology
of chordates as an antipositive curvature inwards may
perhaps be understood as a less drastic equivalence.
New centers defined through meeting of units ...?
It's from eukaryotic cells that multicellular organisms develop
and it's eukaryotic cells that develop sexual cell divisions.
(It's said too that sexual reproduction are initiated at changes
in environment among organisms that have both alternatives.).
One could imagine that in a certain step a duplication
of DNA could occur, leading to meios, a halving as a polarization
← → at cell division,
which the other way around gives the opposite meeting between two
complementary units, defining a new, more highdimensional center:
→ ←.(Also a kind
of inversion of directions, here in the process of reproduction.)
Yet, sexual reproduction doesn't in itself lead
to multicellular organisms. (A condition is perhaps that the small
difference, assumed here in file Genetics,
between "daughter cells" at simple cell divisions, has
grown far enough to complementary poles that imply and stress the
mutual dependence?)
Geometry in positions of cells
may have been a primary factor, perhaps the most elementary,
behind the evolution of multicellular organisms?
There are species of bacteria
(myxobacteria) that first spread radially outwards, then
turn to grow in inward direction towards certain centers and superpose
these, which through these superpositions become defined as spores
(Bc p. 319). Its an example that indeed seems to illustrate
the views here on opposite directions of fields - and a kind of
fertilization through meeting of opposite poles. There is
already an underlying "we" in these groups, assumed as through some
mutual signals.
Cf. how the very relative
position of single cells decide their development to different parts
of a flower according to theories about plants.
In a field of first cells the one
in the center could have been defined as 0-pole, the furthest out
as anticenter, 00-poles, the opposition connected with a radial
versus circular geometry; the central cell becoming defined by its
position as outward directed and consequently with high differentiating
potential, a condition for a multicellular organism. While cells
at anticenter remain a multitude of similar cells. Cf. that this
also characterizes ectoderm in multicellular organisms and cells
from the animal pole at "exogastrulation"
(No. 4).
Now, as far as is known, the evolution of
multicellular organisms from unicellular ones seems to have taken
billions (?) of years. Then we have to count on three historical
phases: 1) cell divisions and "radial" spread of the unicellular
individuals over the surface of the Earth = divergence, 2) pole
exchange to inward direction, 3) convergence leading to meetings
with other cell groups. If so, it would be a parallel on another
time-scale to what is mentioned about the myxobacteria above.
We may compare the spread
of human beings in small groups of kinfolks and clans out over the
world and at certain densities a turn to convergence and confrontations
with other kinfolks and clans, stepwise leading to more hierarchical
structures and superposed organization of societies...
[Level development
One suggested view
in the dimension model here is that a condition for level development
is a meeting between equivalent units (of 5-dimensional types)
with centers as 0-poles, whose mutual relation thus becomes opposite,
defining new more complex centers.
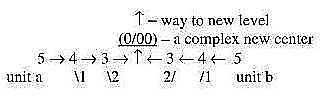 Fig
Ev-7 Fig
Ev-7
The contrast to such level development would be just repeated
propagation:
5 → 4 →
3 → 2 →
1 → 0/00,
as propagation of a single 5-dimensional unit.] A deeper center at start...?
Another hypothesis is the thought that the cells
at divisions of unicellular individuals didn't get quite separated
and retained a closer connection to one another on this fundamental
level that concerns the inner of the cell with relative negative
charge.
The same thing could be described as a cell division
that has reached a deeper level, e.g. the field level of d-degree
4 instead of earlier only of d-degree 3.
It should imply a character of what is called
neoteny, a branch of evolution starting from an earlier embryological
stage of an individual.
The field level - and strength of integrating
0-pole of d-degree 4 in terms of the model - is obviously incorporated
to an internal property in multicellular organisms.
Relative amounts of proteins...?
In fundamental aspects the cell can be regarded as inversion
of an atom. The relative negative charge inside membranes is mainly
given by the proteins, which also make up the radial structure in
cytoplasm and cell membranes with origin from center.
As radial they are principally unlimited outwards - in opposition
to the closed circular component of lipid membranes. (Cf. also the
polarity the FA FG forces
outwards / inwards.)
Thus, if the protein production
became dominating over the one of lipid membranes, suddenly or not,
it could have implied a new degree of communication and transports
between cells - as materialization of field lines.
The clear demarcation of a center
through a nuclear membrane in eukaryotes could perhaps been a factor
in such a change (?).
Light
with its double character of particles and waves could be regarded
as a ground plan for the opposition unicellular — multicellular
organisms: in inward direction appearing as particles, to compare
with individual, separate quanta as cells, in outward direction
appearing as waves, corresponding to the primary, uniting, wavy
proteins on the field level of multicellular organisms.
(What should in that case
among cells correspond to the certain conditions within quantum
mechanics that gives these opposite aspects on photons?)
4. Classification in 1-, 2-, 3-layer animals,
step 2 ← 1:
The division of animals according to number of
tissue layers concerns dimensionally d-degree 2 in the sense
that tissues represent cell contacts forming surfaces. We get the
number chain 1 - 2 - 3 as outer poles of d-degree 2 and 1 and from
1 in the dimension chain. (Cf. that the 1-layer animals according
one theory originates from 2-layer ones.)
The complementary poles 2a 2b out of d-degree
2 in the dimension model are in elementary geometrical terms defined
as outside/inside and/or
convex/concave.
1- to 2-layer species become
more or less bowl-shaped around the gastric cavity in their outer
design.
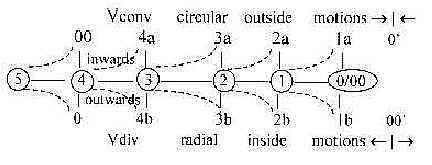 Fig
Ev-8 Fig
Ev-8
(The term species here used without regard to the
scientists classification of levels into kingdoms, phyla, classes,
orders, families, genus, species.)
There have been separate opinions between scientists
on how to classify "1-layer" organisms as sponges since they also
have an inner layer of cells, however more individual, not quite
united to a tissue. They could be called 1.5-layer organisms. The
same regards true Diploblastica, "2-layer" animals as Cnidarians
(hydras, e.g. jellyfishes and corals), which have an extracellular
layer between its two main tissues that can include spread cells,
thus could be called 2.5-layer species.
With reference to the figure
above and "outer poles" as partial structures of each d-degree in
the model such confusion or halfway steps are natural.
About differentiation of organs besides
the gastrula the 1-layer species have only spicules as a kind of
skeleton in the layer of extracellular material besides spread individual
cells inside outer tissue. Among these however a kind of contractile
cells appear as precursors to muscles.
(About forms of spicules
as basis for classification, see file Skeleton.)
2-layer species get further nervous and muscle
functions but hardly as separated organs, only as a differentiation
between single sensory cells besides muscle cells in the same epithelium
(of d-degree 2) and ganglia that innervate the muscle cells.
First with 3-layer species, where the intermediate
layer forms a real 3rd tissue of cells, 3-dimensional
organs develop. Organs in the level
chain as d-degree 3.
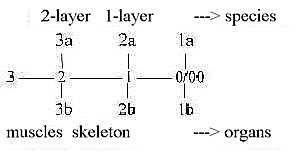
In the level chain of organs
we have regarded skeleton defined in d-degree step 2 ←
1 and muscles in step 3 ← 2. Thus, there
is a natural correspondence between the classification of species
and of organs, especially when counting with the outer poles in
different d-degrees in the model.
 Fig
Ev-9 Fig
Ev-9
(About d-degrees as numbers it could be added
that with a 3rd tissue layer and development of a
front back axis in more developed animals the intermediate
coelom material gets differentiated into 2-3 sections.)
The gastric cavity (the stomach) gets stepwise
differentiated from 1.5- to 2.5-layer species and within these groups.
The step from radial inflows
in sponges to one-way directed inflow (the original embryonic mouth)
in 2.5-layer classes are one example. (It's a drastic change similar
to the one between the circular blastula and definition of a coordinate
A-V-axis in embryology
of chordates.)
The cavity in 1.5-layer
animals is only a sac with inside and outside - as outer poles of
d-degree 2. In 2.5-layer animals as hydromedusae the cavity
is differentiated in a radial canal and a ring canal (Ez p. 62),
the geometrical poles 3b and 3a in our model.
There are however transitional forms in shapes
of the inflow canals: In 1.5-layer sponges the complexity
of the inflow canals increases from straight radial (ascon
type) inwards to angled with side pores in membranes that have become
wavy in convex concave bows (sycon type) to a branched
network of canals with widenings to globular chambers in the epithelium
(leucon type) with flagella of the cells converging
inwards (Ez p. 52, 56), i.e. a more 3-dimensional structure.
It can be compared with how glands
develop in chordates.)
Cf. that these shapes of
canal systems in structures geometrically correspond to d-degree
steps 4 → 3→
in the dimension model, see No. 7 below about "Directions
".
In 2.5-layer species the gastric cavity
becomes divided through walls, "septa", as a stepwise materialization
of the inward direction. (According to reference Fc p.113 they
radiate outwards from center of the gastrula, but according
to the figures on the same page rather from the periphery inwards.)
While hydras lack septa, jellyfishes have partial septa, corals
whole septa.
Numbers of septa are essential in the classifying
system. Septa in the extinct tetracorals were shaped
in 4 steps through three angle steps from a "vertical" axis which
gives number of septa 2 6 10 14 (Fc p.113); compare
number of electrons in s p d f orbitals, intervals
in the 2x2-chain behind the periodic system.
Septa in stony corals (most corals of today) are typically
"6-radiant", have 12 primary septa.
Another differentiation occurs in muscle
functions on septa of 2.5-layer corals (Ez p. 75),
which have "radial" muscles on one side of the septa walls
with divergent, protracting function, and longitudinal muscles mouth
- foot with contracting function along the vertical axis on the
other side. Hence, a polarization both of
coordinate axes to 90°, assumed angle step in d-degree
4 to 3, and of elementary directions outwards/inwards
in function as well as on wall sides, poles of d-degree 2.
Cf. d-degree 3 in the model with outer poles 4b 4a of d-degree
4, Direction, and muscles referred to d-degree step 3 - 2 in the
dimension chain
of organs.
Other features separating these tissue classes
concerns capacity of external locomotion, forming of colonies and
symmetry, see No. 7 below.
5. Coelom —
differentiations, step 3 ← →
2:
Coelom from mesoderm regards the 3rd
real tissue layer (Triploblastica). Within the class of 3-layer
animals the differentiation of this coelom is obviously as said
above a question of polarizations Mass Space, thats out of d-degree
step 3 - 2 in our model. It implies steps from species as Acoela
with coelom as a whole mass to species with splits in coelom (Schizocoela)
to species with coelom divided in outer and inner layers (d-degree
2) with real secondary body cavity (Enterocoela). We have
the inward direction of gravitation (FG) connected
with the property Mass, the outward acceleration force (FA)
connected with Space in macrocosm as well as here, however, in a
reversed relation: inner space, surrounding mass. (Cf. the
cell regarded as inversion of an atom.)
One can describe these
polarizations either in terms of mass space, physical poles 3a
3b out of step 3 2, or just as a step from mesoderm as a volume
(d-degree 3 in our model) to mesoderm as surface layers (d-degree
2).
6. Deuterostomia - Protostomia, step 4 ←
3:
Some characteristic polarities, although not general
in all respects:
|
Protostomia
- nervous system ventrally
- heart dorsally
- archenteron becomes mouth
- mesenchyme from ectoderm (outer layer)
- exoskeleton
- schizocoel common
- spiral cleavage of egg often
- segmentation of body typical
|
Deuterostomia
- nervous system dorsally
- heart ventrally
- archenteron becomes anus
- mesenchyme from endoderm (inner
layer)
- endoskeleton
- enterocoel (real body cavity)
- never spiral cleavage
- not segmentation
|
As said in first paragraphs above this opposition
concerns directions, d-degree 4 and poles 4a and 4b, inward/outward
directions in our model:
- partly inwards - outwards from anticenter
- center poles (ectoderm endoderm) in such things as creation of
skeleton, (exoskeleton versus endoskeleton) and mesenchyme,
- partly in positions of first neural
and nutrition systems in relation to the two coordinate axes Animal
Vegetative poles (A-V) and Front Back (F B), in file Embryology
interpreted as corresponding to d-degrees 4 and 3.)
The complementary polarity of d-degree 4 here implies
also a further "centralization" of separate functions to organs,
representing d-degree 3 in the level chain.
As for coelom in preceding paragraph, its enterocoel
that implies a real secondary body cavity, internal secondary space.
Again, we have the fundamental opposition Mass Space:
Space as such, representing
divergence from 0-pole, here separating mass to tissues in both
directions, seems as a factor behind the domination of outward direction
in Deuterostomia, the properties that lead further to chordates.
Mass of less split coelom
represent convergence and inward direction, thus connected with
Protostomia, including arthropods, the insects.
Why are insects so small and elephants so big?!
Why this difference between enormous amounts of eggs and only a
few? It seems as if it could have its origin in this opposition
between contraction from the 00-pole and divergence from the 0-pole.
Further there were the differences
- in egg division: Protostomia often spiral
cleavage, Deuterostomia never.
- segmentation: typical for Protostomia,
in Deuterostomia no segmentation.
Spiral cleavage of first egg occurs among
Protostomia in many groups of Schizocoela. It implies a kind
of rotation, assumed as motional moment in d-degree 3 in our model.
In Deuterostomia
first cleavages of eggs are "bisymmetrical", vertical and horizontal,
which means along crossing orthogonal coordinate axes that define
a center. In such egg cleavage all 4 to 8 cells have principally
direct contact with the center, the origin. (Cf. stem cells that
separated can develop to whole individuals.)
Thus, the opposite kind
of egg cleavage reflects also this polarity between center - with
"radii" - and anticenter as circular rotation.
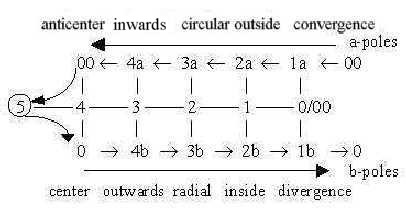
It corresponds to a view
on the dimension model where a-poles of all d-degrees derive from
end of the dimension chain, b-poles from its start:
Fig
Ev-10
Spiral cleavage leads to early differentiated cells
in opposition to the radial and bisymmetric one where the daughter
cells keep a higher potential of differentiation.
Compare perhaps that the
first egg cell in mollusks (Protostomia) undergoes several internal
polarizations in the cytoplasm before the total cell cleavage, while
such polarizations in Deuterostomia as lancelets or frogs (batrachians)
are only one or two.
Segmentation as a division in more or less
equal parts of the body implies a division along the Front Back
axis, the axis which typically gets developed in 3-layer animals.
Its also the axis we have seen as representing d-degree 3 in embryology
of chordates.
Segmentation is typical for big groups of Schizocoela
Protostomia such as ringed worms (Annelids) and Arthropods.
Even mollusks are believed to come from originally segmented forms.
Organs that in segmentation are repeated in each
segment are shells - the exoskeleton, extremities as some kind of
legs, a pair in each segment, nervous ganglia and muscles. That
is organs mainly created from outside inwards the characteristic
direction of Protostomia. ("Superficial" egg cleavage - on the surface,
d-degree 2 - in the class arthropods seems as a connected feature.)
The alimentary canal however from vegetative 0-pole runs through
the length of the body unsegmented. Cf. threadlike colonies of unicellular
organisms that sometimes have a canal straight along the thread
with shared protoplasm.
It has been said somewhere in
a physical context (no reference here available) that convergent
vectors (Vconv) dont reach the center. If so, the segmentation
of worms could be one example!
Why
the multitudes of Masses in macrocosm and the unity of expanding
Space? Perhaps the density of divergent vectors from a center
(not identified or defined) hampers and overcomes the convergent
vectors "half ways" ? Compare Vdiv, the FA-force,
with what is called "dark matter" and estimations of its relative
strength or overweight in relation to mass and the FG-force,
(70 % to 30 % or other similar figures).
Some features of segmentation appear also among
Deuterostomia as chordates, e.g. in muscles of the alimentary canal,
in the straight abdominal muscle of humans, in our notochord and
in bladders on the neural tube in its embryological development.
Segmentation on the level of organs corresponds
to a similar feature on the level of tissues: cells in the outer
tissue (ectoderm, the skin) are more or less equal, while cells
in endoderm have a high potential for differentiation. Compare about
exogastrulation
in file Embryology concerning Deuterostomia: isolated tissue
material at animal pole doesnt differentiate while that from invaginated
ventral pole can develop rather much of organs.
Hence, there is a quantification and multitude
as repetition from outside (the quantifying 00-pole), a continuum
and unity from inside (the quantified 0-pole ). In the dimension
model the force from 00-pole is the primary polarizing one, the
pole from 0-pole the primary integrating one; a relation quantification
continuum related to the one between particles and waves.
Standing waves, longitudinal and/or
transversal, could to a certain degree illustrate segmentation -
as it seems expressed in motions of worms!
 Fig
Ev-11 Fig
Ev-11
A wave and its reflection between two borders correspond
to opposite directions in the dimension chain, the reflection implying
a pole exchange and a quantification. If the illustration is more
than a metaphor, why should the double direction be typical for
Schizocoela Protostomia? The reflected wave as "inwards" along the
F B axis in this group eventually stronger than in the other?
Its said that stationary waves
appear when "the force is not in phase with the velocity".
However
to interpret and apply that statement, one could speculate about
phase displacements of waves as a differentiating factor between
animal groups.
Such phase displacements may correspond to half
steps in the dimension model here, a displacement between a d-degree
and the jump or d-degree step: border interval. Cf. first figure
in this file, differentiation through primary steps.
Examples are the relation between vertebrae
and spinal cord in humans, also development of extremities from
the web between fin bones in fishes.
The opposition plants animals, if expressed in
terms of standing waves could be:
- Plants: longitudinal waves, open ends.
- Animals: transversal waves, closed ends. (Source Wikipedia,
Standing wave.)
Segments can during embryological development and
historical evolution get differentiated functions and be packed
together. In the Protostomia class crawfish (crayfish) the 5 front
segments develop to the head. Note once again number 5! Its about
the same when it concerns the human brain. It seems again to be
a question of positions, the location of cells along coordinate
axes as vectors, here the back - front vector.
Protostomia includes a manifold of groups, Deuterostomia
only a few with Chordate as a big one. Thus, the very number of
groups could be seen as an expression for the polarity manifolds
unity, the 00- versus 0-pole, and the dominating opposition of directions
between Proto- and Deuterostomia.
7. Directions of evolution?
Since all animal phyla of multicellular organisms
seem to have been differentiated already in Precambrian times (Ez),
it make it difficult to establish a direction in time between them.
Among the bases for classifications above only
the one between Protostomia and Deuterostomia represent more clearly
a complementary polarity of poles of the same d-degree in the dimension
model. The other divisions are more as d-degree steps, to or from
more complex forms. To or from? It's easy to be cheated by the clear
evolution within phyla, as the steps from water to land living animals
and more built-in embryos. We can remember that some scientists
regard 1-layer animals as originating from 2-layer ones.
Direction from simple to complex forms is not unambiguous.
In a dimension chain the direction outwards
lower degrees implies increasing differentiation as increasing numbers
of polarizations. This could be translated to steps towards more
simple forms. However, with the differentiations built-in the evolution
goes towards more complex forms. 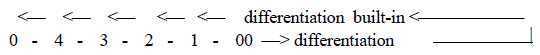
Fig
Ev-12-182-2
A fundamental tone includes all its overtones. A highdimensional
cell or tissue is the one with high potential for development -
like stem cells in relation to specialized ones. In the
embryological development first stages are naturally most high dimensional
in this sense, most "simple" stages as blastula and the
two wall bladder in chordates resembling the 1- to 2-layer organisms.
There are features of these 1- to 2-layer animals
that connect them with highest d-degrees in the dimension chain
and could point to an origin from these steps 5 - 4 - 3.
They are often stuck to
the sea bed or other solid surface or have such stages in their
development. External, "endogenous" locomotion is not
developed.
Density is a factor too (the physical quantity
proposed in step 5 → 4 These
1- to 2-layer classes are often building colonies as the unicellular
ones. In colonies the external relations are still structural, individualization
only partial. (Certain 2-layer hydroids have
the individuals on a common "stalk" where they get mutually differentiated
in function.). Thus features from the further functional differentiation
into organs in 3-layer classes appear already on these elementary
tissue levels.
The symmetry (see below) of 1-2-layer classes
are mostly radial or biradial symmetric, like 5- to 4-merous diploblastica
among plants.
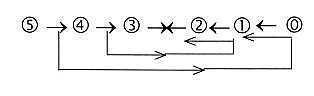
One aspect on this ambiguity may be illustrated by the loop version
of the dimension chain: d-degrees from higher steps debranched and
meeting the other way around in synthesizing direction
inwards:
 Fig
Ev-13 Fig
Ev-13
(It's possible to see some structural relationships
between corresponding steps outwards - inwards in this the loop
version of a dimension chain:
- Naturally the egg with the whole organism (as between d-degree
5 and 0/00).
- The above mentioned inversion of the blastula in a species
of 1-layer sponges and differentiations of directions in Proto-/Deuterostomia.
(4 - 1)
- The septa (developed inwards) in many 2-layer organisms and
of coelom (outward from archenteron) in 3-layer animals:
Fig
Ev-14-179-3x )
In the history of evolution the direction may appear
to be mainly towards "invelopment".Yet, in other respects it seems
possible to suppose simpler animal forms as debranched or reduced
variants of species from more complex levels in the evolution.
5 →>
4 →>3
→>
.......
\
1 \ 2 \
2,5
A more acceptable interpretation in accordance
with the figure is probably to see the high dimensional potential
of an embryo going stepwise further, implying stepwise more involvement
towards more complex forms the other way around: Hence, an evolution
that stops of some reason (e.g. complexity of DNA?), earlier or
later:
5 →>4 →>3 →>2.......3 ←
2 ←
1 ←
0
5 →>4 →>3......................2 ←
1 ←
0
5 →>4.................................... 1
←
0
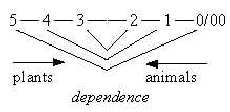
Another aspect concerns the fundamental polarity between plants
and animals. They represent opposite directions
in the dimension chain.
|
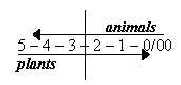
Fig Ev-15
|
 Fig
Ev-16 Fig
Ev-16 |
In essential functions the animals in their involvement represent
the "a-poles" versus plants as "b-poles" in
a dimension chain, "motions to each other", outside in
eating, circular versus radial structure and inward direction embryologically
as in their dependence.
In symmetries the opposition in directions seems clear:
While symmetry in flowers of plants
generally decreases from 2-cotyledons to 1-cotyledons, from 5- and
4-merous plans to 3-merous plans, the polarizations in animals into
coordinate axes become coupled with increasing "invelopment" inwards,
with increasing number of tissue layers:
- 1-layer animals are radial-symmetrical.
- 2-layer animals are radial-symmetrical
- or bilateral symmetrical in certain cases.
- 3-layer animals are bilateral
symmetric.
In the embryological
development the steps from the "vertical" axis Animal-Vegetative
poles to the axis Front Back to the Right-Left axes has been interpreted
as a process through 4th to 3rd to 2nd
d-degree. In numbers of axes corresponding to 1 - 2 - 3.
With only the V-A-axis defined, the symmetry becomes
principally radial, and 1- to 2-layer animals retain thus this main
axis of the invagination gastrula.
With development of the
F-B-axis in 3-layer animals the symmetry becomes bilateral. The
symmetry axes can be described as stepwise "crystallized"
and decreasing towards more "inveloped" animals:
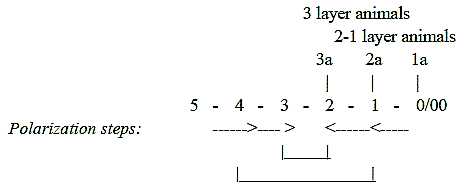
Fig
Ev-17-183-1
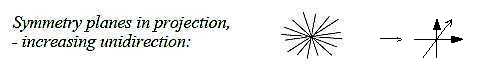
Fig
Ev-18-183-2
(The fundamental reason why the symmetry decreases
with new coordinate axes is naturally that the axes are defined
by complementary poles).
The step from radial to bilateral symmetries resembles the one
in atoms from s-
to p-orbitals: s-orbitals circularly "all-directed"
versus electrons divided on perpendicular coordinate axes in p-orbitals.(Orbital
numbers given from intervals in steps ←
2 ←
1 ←
0 in the 2x2-chain. It's surely not a coincidence, cf.
numbers of septa in corals above.. .
It can be observed that radial-symmetric 2-(2.5-)layer
species as cnidarians often have a 4-numbered symmetry (or
n x 4) in outer and inner structure as related to 4-merous plants.
A primary group of 3-layer
Deuterostomia as the echinoderms is said to have had a 3-numbered
symmetrical form in Precambrian (Fc p. 121), hence a decreased
symmetry towards more "inveloped" forms in agreement with the views
above. Their radial, 5-numbered symmetry of today is a secondary
feature, superposed their bilateral symmetry on their larval stage
(Ez). Compare number 5 in the superposed chain 9 7 5
3 1, halved orbital numbers as intervals in the 2x2-chain
behind the periodic system:
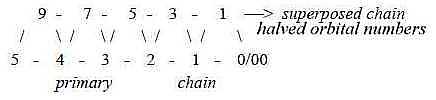
Fig
Ev-19-192x
The 3rd coordinate axis right left
(R-L), the bilateral one, shows many signs to also develop towards
asymmetry, as out of complementary poles:
Examples are for instance
the dominance of the claw on one side in some species of crayfishes
and their lateral motional direction, the increased growth of one
tooth on one side in toothed whales, the differences between right
and left cerebral hemispheres in humans as well as the asymmetric
positions of inner organs and different division of lung lobes.
Etceteras. Birds retain the right arc of aorta, mammals the left
one (Kz p. 210).
In mollusks, believed to have developed from an
early bilateral symmetric species, the intestines undergo a rotation
half a turn that leads to a radical asymmetry bilaterally (Ez
p. 259).
In the dimension model
rotation and spiral motions are assumed as the external motions
in structures of d-degree 3 and 2 respectively. We could wonder
if perhaps a slow process of such motions goes on during history
of evolution, which have proceeded unequal length of time within
different species?
About fundamental tones and overtones, an association:
How overtones on a cello string may appear? Development of tissue
layers and specializations (figure from end of file Embryology).
A"Medusa" figure :
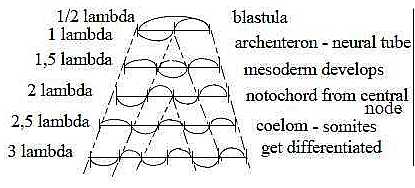
Fig
Ev-20-188
8. Miscellaneous notes:
About secondary polarizations of directions,
numbers and design with associations to the dimension chain:
a) Directions, some examples:
A secondary polarization within the
2-layer phyla cnidarians:
- in hydromedusae do the sex cells and statocysts
derive from ectoderm, the outer layer,
- in jellyfishes the same cells from endoderm,
the inner layer.
3-layer species with 2 shells or valves, polarized
in orthogonal directions:
- Brachiopods, among the oldest, have the
shells along the ventral distal axis V-D.
- Bivalves have the two shells divided right-left,
the R-L-axis.
Both examples illustrate complementary poles
or d-degree steps according to the interpretations here.
Shells of different mollusks illustrating motional
degrees of growth:
- Some have a plane-spiraled shell that illustrates
rotation + pathway motions, a 3-dimensional motion.
- Others have a conical-spiraled shell that
illustrates an extra factor of growth with increasing radius as
a 4-dimensional motion.
- Flagellates: Some has 2 flagella at one end,
others one at each end: a step from 2-way-direction to one-way direction
or the opposite, a depolarization.
b) Numbers of things:
From the aspect of a dimension chain:
5 4 3 2- - 1 0/00, sum 15;
sums of pole values: 10 8 6 4 2.
- Trilobites, old extinct group: 5 pair of legs,
the front pair of which becomes antennae.
- Cuttlefishes (Cephalopods): 10 arms →
8 arms or a multitude of arms (Nautilidae).
- Legs: Spiders 8, insects 6, tetrapods 4, birds,
humans 2, mollusks "1-footed" or "0-footed" (stuck to some solid
surface).
- Body divided: Spiders in 2 parts, insects in
3 parts.
- Wings of insects: 3 pairs first, became 2 pairs.
Mosquitoes and flies: 2 pairs became 1 pair.
- Myriapods (among Arthropods): Diplopods 2 pair
of legs in each segment, Chilopods 1 pair.
Examples of reductions?
- Sea urchins: early species had 15 →
100 shell plates, today 20 ( 5 x 4).
- Fishes: fen rays a multitude →
tetrapods 5 "rays" of toes and fingers.
- Hominids: 5 knobs on the masticating surface
of molars, East apes 4.
An insect as Rhodnius prolixus undergoes
5 stages as a larva with changes of skin before metamorphosis (JB).
Etceteras.
c) Structural design:
Biological shapes on a macro-scale reflect
in many cases structures on underlying levels. According to the
presumptions in the dimension model this depends on the same dimensional
processes reappearing on all levels (cf. fractals).
We could imagine that the structures of light and
of water have "induced" early organic molecules and these further
biological forms: for instance
- the 6-rings of H2O-molecules
in water → rings of carbohydrates
→ leave forms, or
- polarized light with appearing rotation
→ spiraled DNA and proteins.
A fatty acid looks like a simplified form of a
Myriapod.
A ganglioside as a larva that eats leaves, (sketch
after P. Karlson 1976, p. 169):
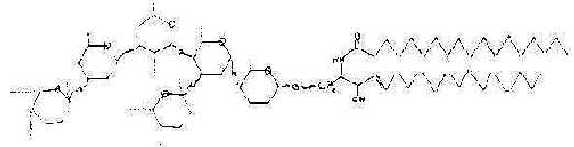
Fig
Ev-21
(Content of this ganglioside, a glycolipid with
aminated carbohydrates: Glucose, Galactose, N-acetylgalactosamine,
N-acetylneuraminate + sphingosine + fatty acids. Mass sum: 1836,
the p/e-quotient, if with uncharged COOH-groups.)
Cf. also the figure here.
- A segmented worm has a macro-form as a circular
magnetic field around an electric cable. Plants
as trees have forms like the magnetic fields around a staff.
- A fish with its fens can resemble the illustration
of a light beam with the fens like the electric and magnetic oscillation
planes perpendicular to the pathway direction.
- A backbone of vertebrae in relation to
the spinal cord illustrates the quantified light beam with phase
displacements.
- All pacing motions have a parallel in this
phase displacement of light propagation.
- Bird wings get a form that resembles motions
of the cell material towards the primitive groove at their embryological
development.
Body forms in the broad outline may become nearly
anything but tend to develop towards the basic dimensional geometries
of d-degree 1 2 3 4, especially among organisms living in
water: needle- or worm-like plane oval spherical star-like
and flower-like or tree-like as divergent vector fields.
Its pointed out by scientists that worm-like forms
exist in many different phyla, e.g. in Mesozoa, Acoela, Pseudocoela,
Schizocoela (annelids - arthropods mollusks), Lophophorata and
Deuterostomia.
Regarded as 1-dimensional
forms, they can be seen from the aspect of one debranched degree
in each step of the dimension chain outwards. Only in the 1- to
2-layer animals these forms are lacking, i.e. where the lengthwise
coordinate axes front-back isnt yet defined.
There are many examples showing that elementary
geometrical forms are more obvious and distinct the closer a structure
is studied in details. (Cf. liver lobes for instance.) In viruses
all round forms are actually polyhedrons.
9. Some general questions:
Characteristic features of classes and species
of organisms are coded in DNA. If now the mutations in genes were
totally haphazard, there wouldnt exist any system in the classifications
of animal groups, no "tree of evolution", no connected properties,
would it?
In that case a multitude of variations and combinations
should exist without any mutual stepwise development between or
within families and classes and it would be without reason to look
for "missing links" ?
If certain genes code for a notochord, other genes
for segmentation, some others for 6 arms, then one thinks that a
segmented chordate with 6 arms should be possible, perhaps even
practical? Flying hominids or trees with brains?
Presumably it shows on the hierarchy of genes,
more general and more specialized, a system that groups properties.
How about complementary genes? Do they exist
in the sense of "complementarity" used in the dimension model? Since
there are proteins with opposite, complementary functions, it seems
reasonable to assume.
And how could such hierarchies develop through
only random mutations and adaptation to an environment?
A fundamental scheme of some kind as the dimensional
model here proposed or the like - seems necessary to presume.
The evolution on different levels, system level,
organ level, tissue level
could also be presumed to follow different
time-scales, which could cause many obscurities in systematics and
relationships.
The big jumps in evolution,
which some biologists talk about, could be imagined as the end of
a dimension or level chain and start of a new? (Or, which becomes
the same, at a certain d-degree in the most fundamental dimension
chain.)
A speculation about the time aspect: If the time-scale
on two different levels correspond to frequencies, and it takes
a certain time for nodes of the two "waves" to coincide, this conjunction
could perhaps imply a decisive mutation? "A certain time" eventually
interpreted as a certain number of generations when the chain of
propagation is regarded as a wave?
Adaptation to the environment is
the other component in the Darwinian view on evolution. However,
the coelacanth, the fish with 4 legs, is said to be "pre-adapted"
(Ez p. 131) to a life on land, i.e. the 4 legs developed
first in the sea before it became practical for a life on land.
Such "pre-adaptation" points sooner to an "endogenous" factor in
the steps of evolution of the here presumed kind, an underlying
dimensional scheme in evolution of genes. (Not "pre-adaptation"
as an anticipation of future life on land.)
(We should perhaps count
on two different kinds of mutations, on one hand the haphazard that
mostly seem to be negative mistakes, on the other such mutations
which represent a given evolutionary scheme of the type dimension
chains in accordance with the model "trial and error"?)
Similarities versus relationships:
In the dimension
model a step of polarization leads to complementary poles
as partial structures). This should imply that animals with complementary
characteristics have a closer relationship than animals showing
similarities but on different levels:
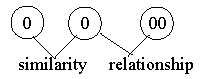 Fig
Ev-22-197-1 Fig
Ev-22-197-1
Protostomia and Deuterostomia could be one example,
prokaryotic and eukaryotic cells another if both derive from Archae
bacteria (now Archae regarded as perhaps a special phyla).
- Relationship could correspond to what biologists
call ramifications in the tree of evolution.
- Similarities appear in what biologists
call convergence, similar features developed in different phyla
as on different levels, "without relationship".
The idea of polarizations
seems inherent in biologists descriptions without being expressly
pronounced.
Just a figure of primary and secondary polarizations:
Fig Ev-23-197-2
An outline
of classifications (freely after Ez and Sb):
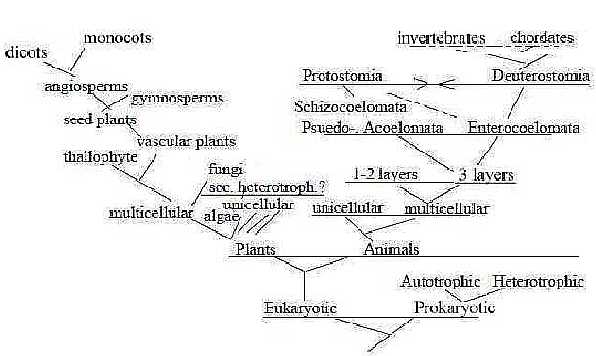
Fig
Ev-24-198-2
Log-scale for times of the Earth; middle ~ 9,65
/ 2 = circa 67000 years ago, a time for
Neanderthals and Homo Sapiens:
Log-scale
9,65 Born of the Earth
9,5 Prokaryotic cells ?
9,3 Eukaryotic cells ?
8,7 The Cambrian most phyla of today
assumed differentiated
8,4 Reptiles. Supercontinent Pangaea
gets split later
7,6 India collides with Asia
6,6 Hominids
5,1 Homo Sapiens
4,0 End of last ice age
3,7 Egypts pyramids
2,2 The industrialism
1,0 ~ New millennium 2000.
To Evolution
- Addition
- Subcellular level
|