|
1. The sense of equilibrium builds on the gravitational
force (FG) as the sense of sight on the electromagnetic
one (FEM). Early forms of chordates as tunicates
had one receptor cell for electromagnetic waves and one "statocyst"
as "gravitational organ" (Kz). In the history of
evolution these organs could be regarded as the "first"
specialized senses for external orientation. (The forces obviously
still more decisive for plants!)
Already in cyclostomes this organ for equilibrium
has developed to sacs with 1-2 ducts, and sharks have the two sacs
saccula and utricle plus the 3 semicircular canals
in three directions as all later species and human beings.
The organs for equilibrium originates
obviously from the lateral line system of fishes, depressed canals
along the sides with sensory hair cells.
Hence, this is a further example of how the environment
(the 00-pole in our model) is successively built-in into an organism
as a center versus the surroundings, a 0-pole: one general principle
view in the dimension model.
A specialization of that lateral line system becomes
the receptors for electric fields among species of fishes, which
implies a step between forces
FG/FA
→> FEM
.
2. The organ of equilibrium becomes divided in 5 structural
parts, differentiated in functions.
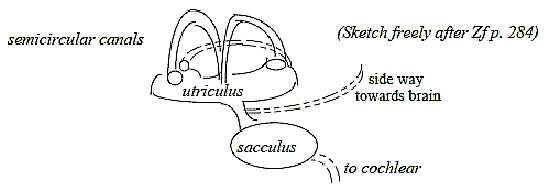
Fig
Eq-1-115
Receptors are hair cells that react mechanically on
the movements of the fluid endolymph in sacs and the 3 ducts at
changes of body and head positions. They are embedded in christae
(called macula in the sacs), one in each duct, one in utricle,
one divided in saccula, hence 5 to 6.
There is a first polarization between sacs and ducts: while the
sacs register static forces, the ducts register changes in velocity
and directions of rotation, also a relation of the kind between
a function and its derivative.
The christae in the sacs react on motions along
gravitational axis up-down and on linear acceleration. Compare our
interpretation
of outward acceleration (FA) as
complementary pole to gravitation (FG) as
inward acceleration. The hair cells in the ducts (the semicircular
canals) react on different kinds of rotation of the head.
This polarization agrees with our interpretation
of external
motions in dimension degree (shortened here d-degree)
step 4 →> 3 of structure:
a 1-dimensional motion developed to a 2-dimensional one (rotation)
in d-degree 3. Simultaneously this opposition implies a step from
the whole body to the part, the head.
(Hence, hair cells in the sacs register linear
acceleration, those in the ducts angular acceleration. This could
also be illustrated with a figure of a dimension chain as angle
steps, a polarity of 180° in d-degree 4, 90° in d-degree
3 as a transition to rotation.)
A functional differentiation between maculae in the two sacs should
reasonably exist but isn't noted in the references here. It's only
said (Mf p. 318) that the function of saccula is less known
but eventually reacts on both linear acceleration and falling -
possibly then both FA and FG
in our terms? It's left as an interesting question.
3. There is however in external form and in arrangement of the
ciliated receptors (hair cells) in the sacs features of partial
polarizations.
In forms the sacs differ as "round"
and "elliptic". An ellipse may be described as a circle,
the center of which has been polarized into two focuses. (Cf. about
the sense of hearing, the "round" and "oval windows"
in cochlea.) The longer coordinate axis of the oval utricle sac
becomes also to a certain degree tangential to the round saccula
sac. The oval sac seems in this sense as an expression of the very
transitional step 4 →>
3, to rotation. Followed by the breaking up of "volumes"
into three perpendicular 2-dimensional planes, designed as "halved
circles" of tube-shaped canals as one expression for d-degree
step 3 →> 2. (2 vertical
ducts, 1 horizontal.)
Cf. 3-dimensional motion
as "translation in 3 dimensions".
4. The arrangement of the receptors in the sacs differ too
according to a figure in a reference (Zf p. 284): in the
"round" saccula they are positioned both vertically and
horizontally with a separate bundle of nerves from each, in the
oval utricle only horizontally, showing on a step towards one-way
direction.
The higher d-degree of maculae in the sacs may
also be seen in the mineral grains of calcium carbonate (the statoconia)
that lie on cilia of the hair cells and through pressure and motions
affect them. This in opposition to only fluid streams that affect
hair cells in the ducts. It's a d-degree step in phase
too of influencing matter.
Further in details: in sacs the relation between
the influencing crystals and cilia is vertical, in the ducts the
influence of fluid streams on cilia is horizontal.
5. The "hairs" of the individual cells are cilia polarized
in two kinds: one big, single kinocilium on each hair cell,
always at one end, and up to 100 smaller stereocilia in parallel
rows. Essential for reaction of the receptor cell is if the stereocilia
are bent towards the single kinocilium or from it.
Here we have both the polarity unity - manifold
of the poles 0 and 00 in our model and simultaneously the directions
outwards - inwards, translated to a linear projection.
In ducts the polarity of directions appears between
vertical and horizontal ducts through opposite arrangements of the
cilia: in vertical ducts the kinocilium is placed outwards from
utricle, in the horizontal duct inwards utricle. Since a reaction
giving a nerve signal is always the result of stereocilia bending
in direction towards the kinocilium, it gives that vertical ducts
in this sense represent outward direction, the horizontal one inward
direction.
Another feature: in the sacs the hair cells give hardly any signals
at ordinary position of the head. In the ducts they have a basic
frequency which varies at different motions of the head. It seems
as an expression for the increasing motional moments towards lower
d-degrees of structure in our model.
6. Signals from hair cells in the 3 ducts correspond to rotation
of head in 3 planes, in at least northwest countries signs for "Yes",
intermediate "Njae" and "No":
- Vertical plane = Front - Back | Dorsal-Ventral axes: posterior
duct, rotation for Yes.
- Vertical plane = Front-Back | Left-Right axes: superior duct:
tilting of head for "Njae",
- Horizontal plane = Dorsal - Ventral |Left - Right axes: horizontal
duct: rotation for No.
It may be noted that the half or decisive 'No'
in these cases include the Left-Right axis, suggested to represent
d-degree 2 according to number of polarization steps in embryos'
development (Embryology,
No. 8): a 'No' also connected with "inhibition",
polarization
step 2-1 in the nervous system.
7. The bundles of nerves become 6 with two from saccula,
one from utricle and one from each duct (Zf p. 284). They
join two and two, which implies 3 polarizations from the aspect
of a main tract.
The bundle from horizontal cells in saccula joins
with the posterior vertical duct: this plane is defined by
the primary two coordinate axes of an embryo,
which we have interpreted as representing d-degree 4 and 3, the
A-V- and F-B-axes. Some geometry guides surely this bundling. Could
the horizontal saccula cells represent linear acceleration outwards,
the vertical ones gravitation? Or the horizontal one both of these
forces, the vertical as secondary one of them? Only guesses.
We can observe in the figure below that the 3 bundles are paired
vertical (V) with horizontal (H) in ducts or relations of macula
and ducts. Saccula as of higher d-degree has both types, a kind
of double-direction.
Posterior duct V-1, superior duct V-2, horizontal
duct H:
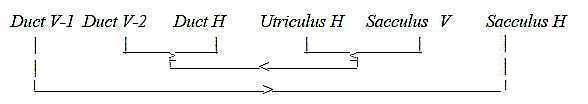
Fig Eq-2-116-2
|