|
First embryological development of an early chordate:
Some features regarded with aspects from the
dimension model.
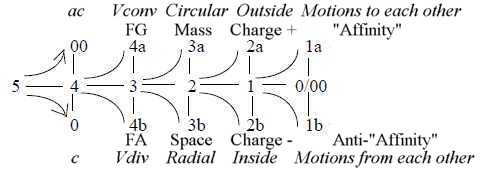
Elementary definitions in geometric and
physical terms in the model with complementary poles in different
dimension degrees (d-degrees):
 Fig
Em-1 Fig
Em-1
Whole 5 - Directions 4 - Volumes 3- Surfaces 2 - Lines
1 - Motions 0/00
Polarizing forces from 00-pole ~ from outside. Binding, integrating
forces from the 0-pole, ~ from inside.
If the dimensional model can be applied, one has
of course to count on a multiple of processing steps at different
levels of underlying and superposed dimension chains more or less
simultaneously.
1. Forms in the first stages of development in an embryo of a lancelet:
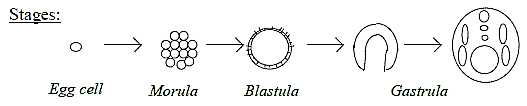
Fig Em-2-11
- First embryo cell undergoes division into a filled ball - a "morula".
During continued cell-growth the filled ball get transformed to
a spherical shell of cells, a "blastula". Its inside room
gets filled with blastocoel liquid.
- A coordinate axis appears defined on the blastula in patterns
of growth between animal pole (A) and vegetative pole (V).
- The following processes - the "gastrulation" - involves
immigration of cell material at the V-pole into the cavity of the
blastula forming a 2-layer bladder and further inner differentiations
into tubes etceteras.
The process can be interpreted in terms of steps
in the dimension model:
2. Aspects on Morula →>
Blastula:
The first step from morula to blastula is rather remarkable and
it may really be asked what guides this transformation. It's mentioned
in another context that outer cells of morula grows faster than
the inner ones and it's is possibly one factor* but cannot explain
why all cells of morula get "center-displaced" to a circumference.
*Cf. in the dimension model velocity
as corresponding to d-degree step 1 →>
0/00, represented in each d-degree step
in the dimension chain outwards.
- It's an expression of Vdiv, of pole 4b in our model, divergence,
a step center to anticenter, (c →>
ac).
- In geometrical terms it's a step 3 →>
2 in d-degrees, from a volume, a 3-dimensional ball, the morula,
to a 2-dimensional sphere, a surface, the one-layer blastula. (Each
higher d-degree in relation to the lower one is a relation of the
type c-ac, ~ 0 to 00: there is an infinity of surfaces in a volume
etc.)
- In physical terms it's simultaneously a polarization of
d-degree 3 into Mass
- Space, as presumed in the dimension model, however inverted
in relation to macrocosm: outer mass, inner space.
(Cf. about the
cell as an inverted atom. In atoms mass is concentrated
to the center, space outside in the shell. Compare too the inversion
of charge over a cell membrane in relation to an atom; negative
charge inside, positive outside.)
- The polarity mass - space gets here the form of a step in phases
between cellular (as "solid") phase (3) and the extracellular
blastocoel liquid (2): note secreted inwards from anticenter.
- A third aspect on these first stages of embryo development is
the relation between the properties Mass and Charge,
presumed in the model to be related as d-degree 3 to 2 (although
level of analysis is regarded as optional). Cf. gravitation,
FG, connected with mass, assumed developed
in step 4 - 3, and the
electromagnetic force, FEM, in step
3 - 2.
One application of this aspect is the mass of
cells of the blastula sphere and the "ion balance" of
the blastocoel liquid, appearing in experiments about exogastrulation,
see below. (Cf. also gravity strongest at the surface of a body
!).
Another application concerns the poles A - V that
develops or appear on the surface of the blastula.
3. Blastula to a 2-layer bladder and A-V-axis:
- In the dimension model the poles out of polarization of d-degree
3 have in elementary geometrical terms been presumed as circular
(3a, with origin in the 00-pole and radial (3b, with origin in the
0-pole). These geometries show up in the growth of cell material:
circular "downwards" from the A-pole towards the V-pole,
radially "upwards" but inwards inner space from the V-pole.
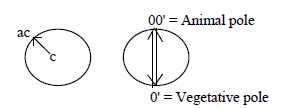 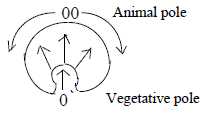
Fig Em-3-12-1, Em-4-13-1
It confirms the identification of the two complementary
poles, and concerns the blastula as a 3-dimensional volume polarized
into mass - space in terms of physical quantities.
Space in macrocosm is interpreted as expanding,
an expression for the outward acceleration force (pole 4b, FA)
as opposite mass connected with gravitation, and inward acceleration
(pole 4a, FG). (With references here to files
about
forces.)
The pattern of growth reminds also of the M-field
around Earth, divergent outwards at the North-pole, convergent
inwards at the South-pole. (N-S-poles known through experiments
to be asymmetric, hence the directions here.)
- The vegetative - animal poles V-A reflect also the physical properties
Mass versus Charge from d-degrees 3 and 2:
Cell material at the animal pole gives a) the
nervous system, operating mainly with electromagnetic signals, b)
the outer surface (d-degree 2) of the embryo, the ectoderm.
Cell material at the vegetative pole gives invaginated
a) archenteron and b) much of the other intestines, hence the nutrition
system that operates with chemicals, in this sense mass.
The two poles represent the two main systems of
the body and its dependence of the environment, its role
as "a half", with inflows from the surroundings.
- Geometrically the step to a 2-layer bladder illustrates further
the polarization of d-degree 2 into outside /
inside (convex/concave), the complementary
poles of d-degree 2 in the dimension model: 2a: ectoderm, 2b: endoderm.
It's said also that long projections of threads
develop from the inside layer of the bladder and draw this inner
layer towards inside of the outer layer. Hence, there is also a
structural development of d-degree 1 in this process in agreement
with the model..
- The remarkable invagination of cell material at V-pole as a main
feature of life implies
a negative or rather antipositive curvature inwards, simply
expressed as surfaces growing faster than the square of radii. (A
necessity if we imagine that there has to be room for a 5-dimensional
unit within a 3-dimensional space!)
In macrocosm there is possibly a negative or rather
antipositive curvature of Space, as expression for the outward acceleration
force FA, and the positive curvature of mass
as expression for gravitation FG inwards.
It should in the dimension model appear as a polarization of Euclidean
geometry in d-degree
step 4 →> 3,
In the multicellular animal the same antipositive
"curvature" gets inward directed, yet as in Space of macrocosm
expanding from V-pole radially into space, while enclosed
through positive curvature of the A-pole.
(In a certain sense it becomes an inversion of
macrocosm also with regard to what has the curvature: V-pole as
representing mass reveals the negative curvature, leading to multilayer
formations in inner space. While the A-pole, as defining the enclosed
space, gets the positive curvature.)
- In the loop version of the dimension model where debranched degrees
from higher steps outwards meet "the other way around",
the step 4→>3 of polarization
into positive / antipositive curvature
leads to the step 2←1,
the polarity inside - outside, forming the 2-layer bladder.
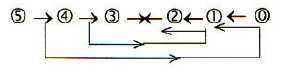 Fig
Em-5 Fig
Em-5
- Simultaneously the processes implies that primary force of divergence,
pole 4b from 0-pole outwards from morula to blastula, in these next
steps is replaced by the complementary force of convergence, pole
4a from 00-pole in agreement with directions in the figure above.
- One general aspect on further steps of gastrulation is that material
at the V-pole as originating from center-pole 0 takes back its position
as center within the material from A-pole, which represents the
anticenter from 00-pole.
What happens with the blastocoel liquid in the two-layer bladder?
It could be presumed that this early liquid contains extra potent
- 'highdimensional' - substances . Formally at least it seems to
land up between outer and inner layer of the skin, between ectoderm
and endoderm. The very rich differentiation of sensory receptor
cells that later develops in the skin could perhaps partially depend
on the blastocoel liquid being an early opposite pole to cell tissue
in the blastula?
The character of V- and A-poles as originating from 0- and 00-
poles and how these are interpreted in the dimension model becomes
affirmed through the experiment called exogastrulation.
4. Exogastrulation:
The experiment implied that the ion milieu was changed in the blastula,
which had the effect that cell material at the vegetative pole didn't
invaginate but nearly separated from that of the animal pole (=
exogastrulation). It turned out that the isolated cell material
from V-pole went through several normal differentiations anyhow,
while the cell material from the A-pole remained a formless hollow
mass.
Interpreted in terms of the dimension model a
center-pole implies outward and hence differentiating directions,
and it has always some kind of anticenter, an outside, a surrounding
with a certain polarizing effect, even if not the appropriate one.
It represents also the integrating force. While a separated anticenter,
the 00-pole, represent the polarizing force and without a center,
something to work upon, remains a hollow mess.
Biologists draw the natural conclusion that the
cell material from vegetative pole is the governing power in embryo
development - possibly depending on a richer or less specialized
cytoplasm in these cells (here we could say a more highdimensional
cytoplasm). Notice that this is an old interpretation from 1970th.
There are surely more knowledge now of how these facts are expressed
biochemically.
5. The neural tube:
The V-pole as the guiding one is further stressed by the condition
for the forming of a neural tube. Not until the invaginated inner
material of the V-pole - the archenteron - has reached up to the
A-pole inside the 2-layer bladder, the central area of cells at
A-pole invaginates to a neural tube. Biologists talk about induction.
It's obviously an expression for a binding force.
In terms of the dimension model the contact between
the poles A-V implies a kind of momentary depolarization, hence
a lowering to a deeper, higher d-degree in which the direction from
the A-pole becomes "vertical" and inwards as in d-degree
4, while it in first stages was circular as in d-degree 3.
This close dependence between A-V-poles, shown
at the very creation of the neural tube, is reasonably one condition
behind the fact that nerve cells through their long axons can find
their special inner organs in the body during the later development.
- In following steps of gastrulation, se below, the growth of material
at A-pole becomes again circular but now along the inside of the
surface, and the growth from V-pole becomes "radial",
expressed in evaginations from archenteron: laterally to coeloms
and upwards to cells for a notochord. Thus in accordance with geometries
of d-degree step 3-2.
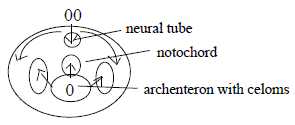 Fig
Em-6-14-1b Fig
Em-6-14-1b
6. The tip over of the bladder:
The two-layer bladder "tips over", which implies that
a second coordinate axis gets defined, the Front - Back one (F -
B). By biologists this has been explained as a result of a "displacement
of the center of gravity" through a change in the storing of
cell material. It sounds as a rather vague explanation and it's
natural to ask: Why this rearrangement of cell material, if so?

Fig Em-7-16
The tip over is a partial rotation, and in our model
rotation is proposed as the 2-dimensional character of external
motions in d-degree 3. That's one reason while we here assume that
the tip over is an expression for the d-degree step 4 →>
3. The connection with changes of gravitational center and mass
- as these concepts are interpreted in the model - agrees with the
same view (gravitation
as a force from pole 4a).
Obviously the development of this new axis Front
- Back is not a result of outer gravity and position or related
to an external source of nourishment but endogen. The same should
be valid for the A-V-axis, the development of which has been assumed
by biologist to depend on a richer cytoplasm at the vegetative pole.
Thus, reference to concepts as gravity and mass reappear here about
the F-B axis.
(Another reason to regard the axes as given from
inside, of endogen nature, is that also unicellular flagellates
include a central axis in direction of flagella.)
If the A-V-axis could be said to represent d-degree
4 or step 5 →> 4, the new
axis Front - Back may be seen as representing d-degree 3 or step
4 →>3. In angle steps as
halvings it's a step 180° →>
90°, which has been assumed in this step in the hypothesis about
dimension chains as angular polarizations.
With the F-B-axis defined the inner, enclosed
volumes turn to the lengthwise, tube-formed growth.
7. Mesoderm - the further development;
- The subsequent process of gastrulation implies the mentioned invagination
of animal pole to the neural tube - and evaginations from the ventral
pole, i.e. from archenteron: an opposition in agreement with our
definitions of inward and outward directions from 00- and 0-poles
respectively.
In general terms the differentiation of inner
V-pole material implies laterally secondary polarizations mass -
space into creation of a true body cavity (enterocoelom) with enclosed
tubes.
- The circular - radial polarity of growth from original A-V-poles
characterizes also the further process as shortly mentioned above.
Material around the A-pole and neural tube invaginates too (inward
direction from anticenter) and spreads circular (from pole 4a to
pole 3a in the model) along inside of ectoderm - while the evaginations
of endoderm from central V-pole have radial and upward direction
and becomes tied off parts. The "vertical" one becomes
the notochord (and in further historic evolution replaced by vertebrae
that comes to enclose the neural tube), and laterally (or in an
angle as ¨45°") to coelom, mesodermal material.
Fig
Em-8(6)-14-1b
With the coelom and tube-formed growth along the F-B-axis the 3rd
coordinate axis Left-Right (L-R) inevitably gets defined.
- One opposition of the type center - anticenter between V- and
A-poles appear in their different contribution to mesoderm: from
A-pole material it's in the form of mesenchyme, i.e. individual,
immigrating cells; from V-pole material it has the form of cell
layers: a difference in d-degrees of structure that shows on the
more highdimensional force of integrating V-pole.
- Another step in d-degrees, 3 →>
2, is illustrated by the first development of coelom from archenteron
in the form of lateral plates: 2-dimensional screens that
grow to surround archenteron as a 3-dimensional volume (and also
notochord and neural tube as volumes).
- A corresponding step in lower d-degrees, 2→>1, can be identified at the animal pole in the creation
of a neural 1-dimensional wall from border of the 2-dimensional
neural plate when this invaginates to the neural tube. A fold that
also defines the central line between right and left sides. On a
tissue level the process can geometrically be described in d-degree
steps 2 →> 1 →>
0, with the last step represented by neural wall material invaginating
to disintegrate into individual mesenchyme cells.
The process as such could further illustrate what
in the model here is the "pole exchange" in d-degree 0/00
of motions: pole 1a, "motions to each other" is shown
by convergence of folds to the neural wall and then inwards. It
defines according to the model a new 0-pole (0') from which we get
outward direction, "motions from each other", as shown
by the divergence of cells from neural crest inside ectoderm.
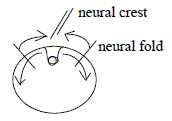 Fig
Em-9-14-1a Fig
Em-9-14-1a
Further development of coelom:
The whole process looks roughly like repetition on the mesodermal
level of steps from underlying, more elementary steps of gastrulation
in 1- to 2-layer organisms. However reversed.
With the conditions of a circular growth from
A-pole and all new material having to enter through invagination
from ventral side, the development of mesodermal coeloms seems to
imply a kind of "pole exchange", a secondary "A-V"-axis
turned upside down relative the original one:
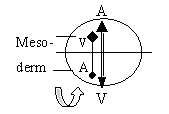 |
→ Upwards
- Ventral pole
→ Downwards - Animal pole
|
Fig Em-10 |
Material nearest first V-pole, "south of
the equator" of a blastula sphere, should at it seems evaginate
to front part of the lateral plates. After a split into 2 layers,
the inner tissue layer thickens and becomes the somites that develop
to striated musculature, shoulder skeleton among other things.
Material "north of the equator", closer
to A-pole on a blastula sphere, should invaginate later and with
this view form the ventral, back part of mesoderm. After split of
the plates the inner layer of these coeloms seems to be the origin
of smooth, visceral muscles.
Regarding directions, the striated muscles from
front part serve outward locomotion, the visceral ones intestines
etc. direction inwards with this functional aspect. A reversed A-V-axis
could explain such data and also why striated muscles grow to an
essential part of body mass.
Below an effort, indeed very rough and approximate,
to illustrate the outgrowth, separation and and split of lateral
plates to coelom:

Fig Em-11-017-2
Geometrically the split of the 2-cell layer of lateral plates implies
other kinds of inversions too in chordates who get a true body cavity:
- Directions outwards/inwards (step
4 →> 3) of layers
get reversed - of natural reasons: Inner layer get the inward direction,
becomes outside of intestines, while outer layer gets the direction
outwards and becomes inside of skin or the abdomen.
- Layers of plates versus cavity can be regarded in terms of the
primary polarity mass - space in d-degree step 3 →>
2 according to the model. Inner central cavity as divergent
space becomes reversed to space as anticenter in relation to all
intestines, which become more or less centralized masses. (Biologists
explain the split of lateral plates as caused by "anti-affinity",
which probably is chemically expressed in some terms of charge (a
property assumed of d-degree 2 in our model). Anti-affinity corresponds
to "motions from each other" in the model, pole 1b, which
implies divergence and thus can redefine space as anticenter.
- The first split of the 2-cell layer of plates (d-degree 2 as
the one of surfaces) to 2 layers can simultaneously illustrate the
polarization in d-degree step 2 →>
1 in the model, into poles 2a - 2b as outside - inside. This
should imply a definition of d-degree 1 according to the model,
which here seems to be nonsense, if not the first rupture between
the layers may be seen as such
Another possibility is to regard the thin, small
"bridge" between front and back part of the plates as
a bit more substantiated expression for 1-dimensional linear connection.
In this case the polarization would concern another direction, the
F - B axis
We have also that longitudinal "ridges"
- as "linear" formations - develop in the neighborhood
of these bridges, ridges which give material to organs for excretion
of fluids (kidneys) and for gonads, the reproductive system; hence
possible to see as the last step 1 →>
0/00 (~5') in our model.
Real development of the proposed steps above is mainly the opposite
to descriptions above.
A special question is why mesoderm gets so
clearly divided lengthwise along the F-B axis in a front and a
back part with only a thin bridge connecting front somites and
back coeloms(In humans marked by the diaphragm.) It's a polarization
to a certain degree complementary too. Do the lateral plates eventually
grow out in 2 opposite directions? It looks as if the imagined
"equator plane" of a blastula stage became more than
virtual. One aspect perhaps: Development of coelom implies
that the Left - Right axis gets defined. Together with the primary
A-V-axis (dorsal - ventral) it defines a plane that divides
the Front-Back axis. Dimensionally it should in d-degree 3 be
d-degree 2 as a potential surface that act as polarizing force.
This third halving implies a mix of the earlier
to complementary poles in two opposite directions, a division
in 4 poles, hence the clear opposition between somites and back
coelom.
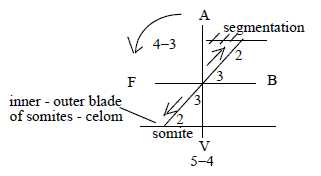
8. Coordinate axes:
Coordinate axes become defined as results of polarizations between
complementary "poles" (the poles also equivalent with
partial structures).
Each new coordinate axis is regarded as polarized
into next one.
 Fig
Em-12-18-2 Fig
Em-12-18-2
The 3 coordinate axes developed as from angle steps between d-degrees:
- First axis A-V defined through d-degree step 5 →>
4, angle step 360°→180°.
- Next axis F-B defined through d-degree step 4 →>
3, angle step 180°→
90°.
- Third axis L-R defined through d-degree step 3→> 2, as an angle step 90°→+/- 45°.
The d-degree steps represent also development of the three layers
of 3-layer organisms:
- A-V-axis the 1-layer stage of the blastula,
- F-B-axis the 2-layer stage of the bladder that tips over,
- L-R-axis the 3-layer stage with developed mesoderm.
With the loop version of the dimension model (figure 4 above)
we get the number of tissue layers connected with angle steps and
coordinate axes in this way:
5 → 4: 1-layer stage: morula
→> blastula: in opposite
direction 1← 0/0
, the 1st axis A-V.
4 → 3: 2-layer stage: blastula
→> bladder: in opposite
direction step 2 ← 1, the
2nd axis F-B.
3 → 2: 3-layer stage: mesoderm
develops: in opposite direction step 3←2,
the 3rd axis L-R.
(A halfway meeting of directions in this 3rd polarity:
90°→ 45°. One
aspect on the approximately bilateral symmetry developed in this
stage?)
s-p-orbitals in electron shells:
The evolution of coordinate axes from the blastula resembles how
the "circular" s-orbitals of electrons in
atomic
shells are followed by the orientation of electrons along
three coordinate axes in the p-orbitals. There is
the similar process from s- to p-orbitals as in the
embryo from 360° to 180° to 90° of polarities in orientation.
In the next, higher d-orbitals,
representing the middle step 3 - 2 in the 2x2-chain behind
the periodic system, there are also electrons divided along axes
in the plane quadrants, i.e. in angles of 45°.
The axes correspond also to cleavages of cells
at first stages of fertilized eggs in many organisms.
The A-V-axis, the first defined, mark as pointed to above
the positions of invaginations, the primary inflows from the
surroundings: 1) inflow of material (cf. Mass at V-pole, archenteron
as first mouth) and 2) sensory inflow (cf. conducted through Charge,
at A-pole). In this sense it represents the cell as "a half"
in relation to its total environment as the other half, as the angle
step 360° to 180° is a halving
F-pole represents a secondary form of animal A-pole
after tip-over of the bladder. Here an invagination occurs in this
type of organisms, Deuterostomia: the secondary mouth opening,
(inward direction from anticenter): an analogy to invagination of
neural tube at first animal pole.
The F-B-axis implies in relation to the A-V-axis
a step from two-way direction to one-way direction mouth —
anus in chordates.
The direction of external motion gets defined,
at least for most fishes and land living species apart from Homo
Erectus, where the original A-V-axis dorsal - ventral give the direction
A → V.
The 3rd L-R-axis gives the approximately bilateral
symmetry that also characterizes many 3-merous
flowers.
The polarizations of mesoderm can be described as more or less parallel
to the primary ones:
- inner versus outer layer of split sideplates analogous to polarization
ectoderm / endoderm,
- somites versus ventral coelom parallel to front - back axis and
- later partial segmentation of e.g. striated muscles parallel to
the lateral axis L-R.
(Paired organs as e.g. kidneys and ovaries seem to be the result
of this 3rd coordinate axis, not otherwise necessary in themselves.
Material from mesoderm takes part in these paired organs.)
9. A summary of dimensional views:
It seems surely too easy to identify the elementary geometrical
forms proposed in the dimension
model in this complex embryonic development. Yet, it's hard
to explain the "invelopments" without a similar model.
We have the polarization Mass - Space from morula
to blastula, defining a surface, the circular versus radial growth
during invagination from V-pole, a polarization outside - inside,
which also implies the long projections between these as of d-degree
1 that draws the layers towards each other. (In d-degrees: Morula
(3) to blastula (2) to projections (1). And behind these processes,
necessary to assume, a field level of forces, guiding in first step
the outward direction, in next step the invagination, equivalent
with directions outwards / inwards. In
all steps and further on expressed in d-degree 0/00
as motions from/to each other.

Fig Em-13
10. Some more general aspects:
a) Affinity:
Why do cells in the blastula keep together? Biologists refer
to "affinity" between the cells and the formation of tubes
is for instance explained as result of an especially strong affinity.
Cracks or splitting of a cell material is referred to as anti-affinity.
However, it's hardly an explanation.
It's naturally another word for acting forces.
These are certainly expressed in some biochemical substances but
here these are assumed as just expressions for and "carrier"
of underlying forces - polarizing/binding
ones - in the same sense as photons are called carriers of the electromagnetic
force.
In the dimension model we have convergence from
anticenter. (Cf. gravitation, strongest at the surface of a body.)
On the chemical level there is the hydrophobic bond, corresponding
to the nuclear force and appearing in cell membranes.
We ought to imagine the atomic forces reappearing
here, "extrapolated" to this multicellular level in reversed
forms. (See further The
Cell.)
It could be observed that the embryological step
morula to blastula have a certain correspondence already on the
elementary molecular level with the step from tetrahedrons to molecular
carbon rings, through sp-hybridization.
All primary physical quantities,
regarded as a dimension chain, should naturally be expected as expressed
and involved in the embryonic differentiation, with density, added
as only conceptual quality in step 5 → 4: forces (d-degree 4), distribution of mass (d-degree
3), ion balance (charge, d-degree 2 according to suggestions here)
as well as distances (d-degree 1) and time (d-degree 0/00
of motions) as in relative velocities of growth.
Invelopment as such could in generalized terms
be the effect of binding forces from higher d-degrees (2←1←0
from 5 → 4 → 3). The ultimate binding force behind the invaginations at
animal and vegetative A-V-poles would be the binding forces from
d-degree 5 between 0 and 00 leading to the creation of a multilayer
organism within first poles.
On superposed levels similar "affinities"
between individual multicellular organisms can form groups that
may function as more or less one single organism, as among cnidarians:
a 40 meter long "worm" of jellyfishes where position
in the row defines their different organic function!
Cf. human clans as well!
The polarizing force, the "anti-affinity", appears very
early, as it's said that first cell division is initiated from cytoplasm
of the fertilized egg cell. One to several divisions occur also
within a first embryo cell of different species before the whole
cell divides.
It's said too that eukaryotic cells, even without
nuclei (artificially picked out), odd enough can divide (one reference: http://www.sciencenews.org/pages/pdfs/data/1996/150-09/15009-16.pdf).
What in membrane or cytoplasm induces this polarization?
In any case a force from anticenter, a 00-pole in our model.
The blocking of genes in DNA that differentiates
activities of cells may also be regarded as a kind of polarization
and occurs through molecules from outside.
(If we regard DNA as a kind of piano and should
translate the 24
codons to two octaves of tones, perhaps the 5 G1-codons
and the 5 C1-codons to the black tangents and the 7 U2-codons
and 7 A2-codons to the white tangents, how should the melody
of e.g. insulin or substance P sound? But DNA is sooner like
a string instrument where blocking defines a tone, than a piano)
One factor behind the fundamental "affinity" that differs
multicellular organisms from unicellular ones could perhaps be identified
in terms of what is called "neoteny": a branch
of evolution starting from a deeper, preceding level of embryonic
development. Translated to the dimension model it would imply a
start from e.g. level of d-degree 4 instead of 3, a still less differentiated
one, where the binding force would be stronger.
Communication over distances in the developed body
- such as nerves finding their special targets or hormones their
receptor cells - is in reality the same kind of mystery as the old
one how gravitation can act over distances in macrocosm. It could
also be seen as an expression of "affinity". As in macrocosm,
it's the forces in development of space that create the distances.
At some first cell divisions the individual cells can develop to
complete organisms and obviously contain all information. Later
cell divisions imply specializations.
Hence, it seems necessary to assume that the division
of a cell at this stage gives daughter cells that in some respect
are complementary. Hardly in the genetic code itself. Perhaps in
which genes that becomes blocked, although pre-decided somewhere
in DNA and rooted in geometries. (Cf. Epigenetics.) Some essential
information seems lacking about how such a differentiation between
daughter cells is realized (at least here in used sources).
Anyhow, such polarities at cell divisions could
be suspected to be one factor behind communication over distances.
With the dimension model the distant connections could be apprehended
in more abstract terms: an inner relation, represented by a higher
d-degree get when polarized "inverted" to outer connections
into next lower d-degree as a new potential, including new directions
and a new distance.
 Fig
Em-14 Fig
Em-14
Another aspect on the question is level chains. It's assumed in
the dimension model that each step in a fundamental first chain
can develop to whole new chains of 2nd order, steps in these to
whole chains of 3rd order etceteras., which leads to a "level
chain". (In the figure below illustrated as simple loops in
each first chain steps.)
 Fig Em-15
Fig Em-15
An original close relation of a d-degree step in a basic chain
can in this way develop to a very far one.
Compare how mRNA is cut on its way to ribosomes:
a way to reestablish the original basic relation in the hierarchy
through cutting off the loops?
b) Processes of design as waves:
Tube-forms as archenteron and neural tube, which also
were explained as results of extra strong affinity between cells,
may be regarded in the same way as in- and evaginations of a cell
membrane in a single cell, cut off to vesicles, serving import and
export of material. (Cf. about the cutting of mRNA above.)
They can be interpreted as the 3-dimensional external
motion attributed to a 2-dimensional structure - the tissue level
- in the dimension model.
With addition of a 4th motional moment, the growth
lengthwise along the front-back axis F-B the forms become "1-dimensional"
canals on a macro-scale.
The wavy character reminds of the problematic turbulence of water
streams where bigger whirls get translated into smaller and smaller
circular whirls: a way to store a surplus of energy?
Another parallel is found in meteorology: how
high- and low-pressure "cells" are tied off from the Rossby
waves around the North Pole. Why are they tied off? It seems necessary
to assume that a certain area on the surface gets defined and demarcated
with the border defined as anticenter, from which convergence leads
to the tied off "loops" - as occurs with neural tube from
neural wall.
Tubes can also get designed another way as from
d-degrees 0/00 inwards 1 and 2 from single
mesenchyme cells when it concerns a fluid as blood.
In the dimension model it's a general hypothesis that the d-degree
of motions increase towards lower d-degree of structure. Internal
structural relations are stepwise released to external motions.
Simplified: The opposition waves versus particles are seen as lower
d-degrees of structure in relation to higher ones.
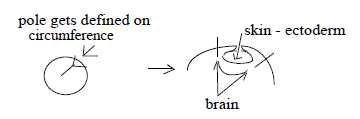
Segmentation could also be regarded in terms of waves.
As previously mentioned the inner front part of somites gets segmented.
Segmentation in the form of transverse bands takes also place of
the neural tube during its embryonic development. That's quantification
in rows of similar units, a linear repetition in first phase, which
later may be more or less differentiated. (Cf. Evolution
and segmented animals.)
It should be noted that segmentation concerns
typically front and dorsal part of the embryo, representing the
earlier A-pole (00), corresponding to primary polarizing force in
the dimension model.
This feature of segmentation has the character
of longitudinal L-waves lengthwise:
→ 0 ←
00 → 0 ←
00 → 0 ←...
, a variation in density, assumed as first physical quantity in
the model in step 5→4.
Such linear, 1-dimensional L-waves are suspected on the field level,
in d-degree 4 (as in gravitational waves if they exist). Cf. that
they in typical, segmented species demarcate whole units of essential
organs.
In synthesizing directions, inwards from end of
a dimension chain, motions to and from each other, it becomes one
way to build 1-dimensional lines. On a molecular level it could
be compared with the way globular proteins or units of collagen
line up after one another.
Neural tube - vertebrae give in their relation another
example of how elementary waves can be applied as aspect on design:
The vertebrae come to enclose the spinal cord
as magnetic field lines surround an electric conductor.
There is a phase displacement too between vertebrae and segments
of neural tube as between M- and E-components in an electromagnetic
light wave, in this case a transversal T-wave.
First notochord and the neural tube derive from
opposite poles V-A in the embryo, which could underline the complementary
polarity in this later design vertebrae - neural tube.
A general view could probably be to see the whole embryonic development
as materialized processes, substantiated motional structures, or
formulated as "captured internalized waves", whatever
to mean by that.
c) Geometry:
It seems obvious that the position of cells in the blastula decides
their later roles and functions. It could be taken as evidence for
geometry as the guiding principle.
A general aspect from the dimension model is also steps towards
more and more differentiations of directions, i.e. towards increasingly
specialized ones. Cf. about level chains above.
The c-ac polarity center - anticenter appears not
only in the first phases of whole embryo and the polarity along
diametrical axes. Such polarizations occur obviously also within
regions on the tissue surface, i.e. in d-degree 2
Best known example seems to be the area around
the animal pole, where the neural plate gets defined as center and
surrounding circles of cells immigrate and come to differentiate
in 8-10 separate cell types (Kz p. 116) - as from circles
of different radii from the center (?). They include more peripheral
types of nerve cells, glial cells that surround nerve cells, cells
for membrane around the brain, cells for teeth, for cartilage and
bone in cranium...

Fig
Em-16-22-2
How position of a cell in morula - or generation number? - is translated
to its position at surface of the blastula has perhaps been clarified
during later decades (?) - but what decides the angle, demarcating
the neural wall as border between immigrating cells and those remaining
ectoderm?
(Material that becomes mesoderm from archenteron
is also anticentric around the vegetative pole.)
Induction, the term biologists use for certain processes,
includes also special geometrical features in design. It's the same
term used for how E- and M-factors influence each other in a magnetic
field. Thus, it concerns how complementary biological poles influence
each other, expressed with views in the dimension model.
One essential example is the mentioned invagination
of a certain area around the animal A-pole to a neural tube through
induction by the invaginated vegetative V-pole when it reaches the
inner "ceiling" of the early 2-layer bladder at A-pole.
It represents a partial or momentary depolarization to higher d-degree
of primary A-V-poles, where the upward direction of V-pole induces
downward direction from A-pole. The induction should go via the
higher d-degree as binding force and underlying, deeper level -
as d-degree 4 in relation to poles 4b-4a of d-degree 3.
More or less simultaneously (?) the "ceiling"
cells of archenteron from V-pole develop to the germ for a chord
or notochord, also straight upwards. It could point to induction
as eventually mutual.
There is however something more in this process, perhaps from
mutual induction. Vertebrae come to enclose enclose the neural tube.
Material with origin from V-pole, principally radial and outward
directed, become anticenter around material from A-pole, principally
inward directed, which thus gets the role of a center.
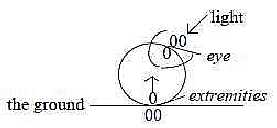
Another example is how the eye lens is formed:
it requires the meeting between the eye cup from inside the brain
with ectoderm from outside, which through this induction gets formed
to the lens.
 Fig
Em-17-23-2 Fig
Em-17-23-2
In these and other cases it is not only a question of chemistry
but of geometry, a center - anticenter polarity in the design and
a "pole exchange". Divergence from some centers inside,
convergence from outside leads to a structure where inner material
become anticenter around outer material. Pole exchange" is
in the model assumed to occur in last d-degree of motions 0/00,
and a step 1 - 0/00 in either direction
should be involved - as it is by definition in every d-degree step
in a dimensions chain.
A question is how the surface area for invagination in these examples
is defined, through a certain solid angle of virtual radii from
inside?
Other examples of a similar geometry in the human body:
- Hypophysis from a pharyngeal pouch that come to enclose nerve
material from the brain.
- Adrenal glands with the bark (ac) from intestines (vegetative
pole, ~ 0) and their marrow (c) from the nerve tissue (animal pole,
~ 00).
- Oviducts in relation to ovaries.
- In skeleton ball and socket joints in hips and shoulders.
The aspect on the eye lens and its construction could be extended
to the meeting between the whole organism as 0-pole and direction-specific
environment as 00-pole: new complex centers could be expected defined
at this pole-meeting on the surface of the organism and be built-in
into new structures (as it occurs at the psychological level).
Examples of course are sense organs but maybe also
fin rays, where bottom, ground contact could induce rudiments of legs,
of new skeleton parts under the skin...? (A geometric approach to
the evolution...?)
 Fig
Em-18-24-1 Fig
Em-18-24-1
Gradual incorporation of the 00-pole is a more general
formulation of the corresponding principle (cf. inward direction from
the 00-pole):
- the incorporation of sperm chromosomes into the egg,
- the incorporation of surrounding space in neural tube and archenteron,
- the incorporation of external nourishment,
- on chemical level also the building-in of metal ions (00-poles in
relation to non- metals etceteras.
(Universe as a camel trying to pass through the
needle eye of a cell!)
Number of cells becoming the human, amniotic embryo
after a certain amount of cell divisions is said to be 4 - 5. All
the other become coverings, some with the function to penetrate
into uterus. Apart from the interesting number, it shows obviously
that it is the geometrical position as center that indeed decides
the potential for development. It confirms too what was find in
experiments with exogastrulation described above.
d) Mass - Space:
Masses are built-in into vacant space in macrocosm and "vacant"
space as primary antimatter built-in into atomic structure in microcosm.
We could see gastrulation and multilayer structures as a similar
double process at a size level between micro- and macrocosm.
Surrounding space gets built into cavities as
"negative mass" within closed cell layers in neural tube
and archenteron, coelom etceteras., and these get built-in as positive
mass in the cavities of the embryo.
And as presumed in text about physics atoms "breathe
vacuum" and are depending on surrounding "vacant space"
for maintaining their structure as atoms, so the embryo with archenteron
and neural tube supports itself on its environment.
*(File about the atom in Physics not yet translated.)
A multilayer creature from repeated positive - negative curvature:
 Fig
Em-19-22-1 Fig
Em-19-22-1
A note:
It's easier to visualize the whole embryological process
in d-degrees 3 - 2, and perhaps we should imagine the process as
a stepwise "inversion" of deeper levels through step 3
— 2 to superposed ones?
The cello string - an association:
A speculation, without any knowledge, about vibrations in a cello
string:

Fig Em-20-28-2
Quantification of waves with obscure allusions to biology:
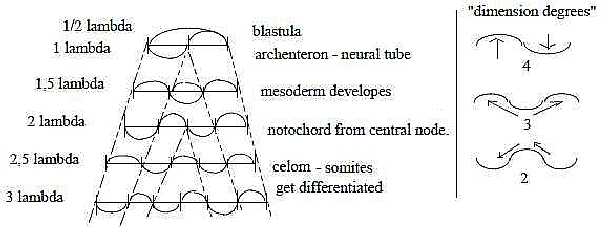 Fig Em-21-28-3 Fig Em-21-28-3
Displacements of side-waves through new vibrations
as "serial inlays" from the middle. With half-steps: in
every second step bellies in the middle, in every second nodes,
witch imply tied off centers.
|